ক্রিকের সেন্ট্রাল ডগমার শেষ ধাপ হলো ট্রান্সলেশন। এই ধাপের জন্য তিনটি জিনিস দরকার হয়। প্রথমত, DNA-এর কপি বা mRNA (ম্যাচিউরড-mRNA)। দ্বিতীয়ত, ট্রান্সলেটর বা অনুবাদক। তৃতীয়ত, প্রোটিন তৈরির যন্ত্র বা রাইবোসোম (Ribosomes)।
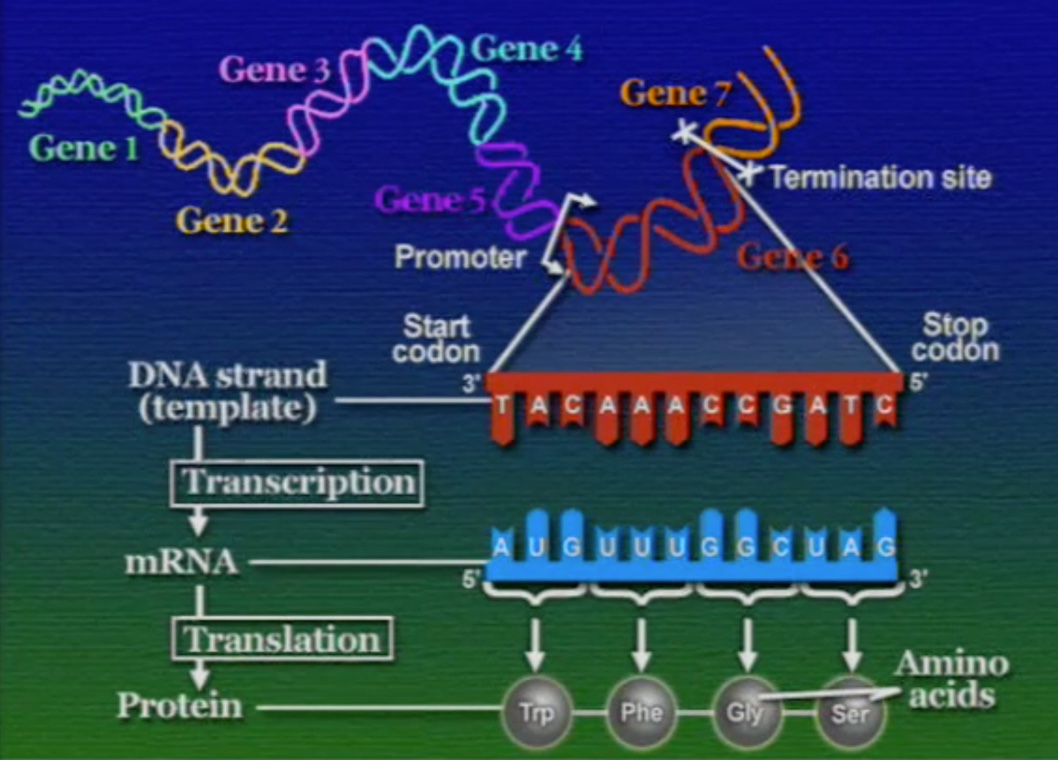
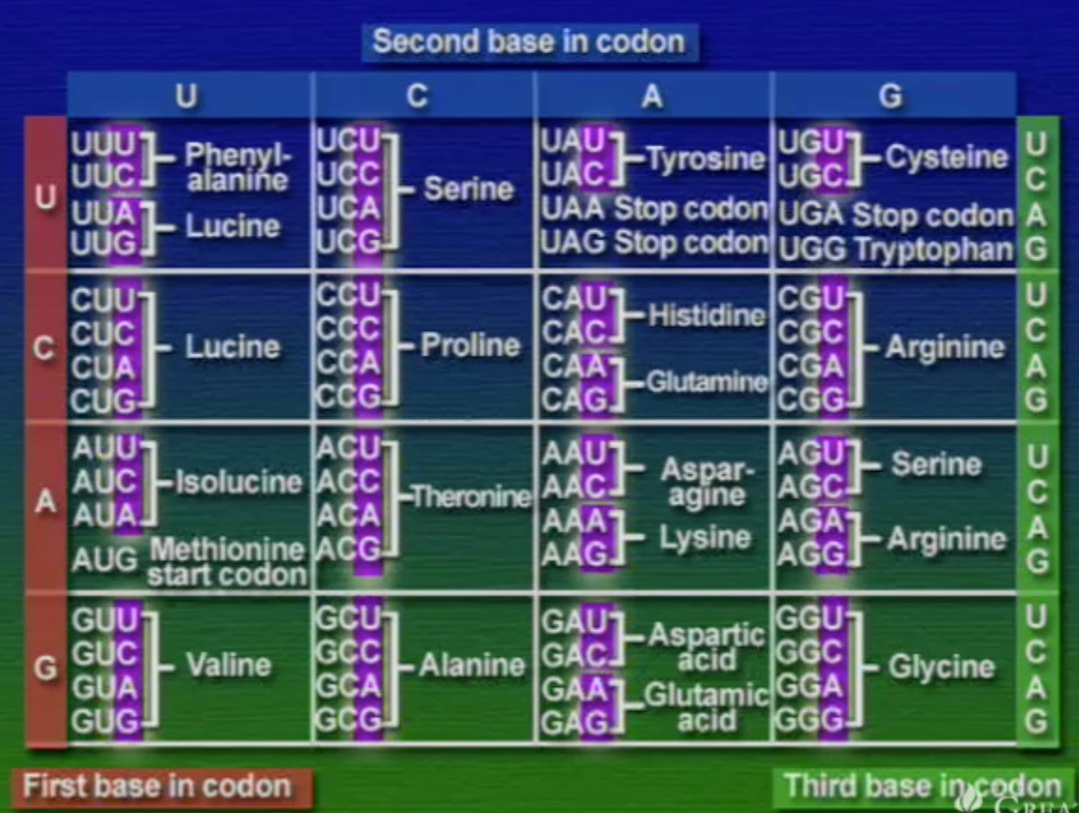
শুরুতে বলে রাখা ভালো, mRNA তৈরি করা হয় DNA-এর কমপ্লিমেন্টারি বা পরিপূরক নিউক্লিয়টাইড জোড়া লাগানোর মাধ্যমে। এই mRNA-এর পরপর তিনটি নিউক্লিয়টাইডযুক্ত ট্রিপলেট কোডিংকে বলে কোডোন।

ইউক্যারিয়টিক কোষে mRNA তৈরি বা ট্রান্সক্রিপশন প্রক্রিয়াটি চলে নিউক্লিয়াসে। অন্যদিকে, ট্রান্সলেশন প্রক্রিয়াটি চলে নিউক্লিয়াসের বাহিরে, কোষের সাইটোপ্লাসোমে (Cytoplasm)। প্রোক্যারিয়টদের নিউক্লিয়াস নেই তাই ওদের ট্রান্সক্রিপশন এবং ট্রান্সলেশন দুটিই চলে সাইটোপ্লাসোমে। আমরা আগে দেখেছি, DNA ও RNA দুটোর ভাষা এক, অর্থাৎ, দুটোই নিউক্লিইক-অ্যাসিড দিয়ে তৈরি। অন্যদিকে, RNA ও প্রোটিন দুটোর ভাষা আলাদা, প্রথমটি নিউক্লিইক-অ্যাসিড আর পরেরটি অ্যামিনো-অ্যাসিড দিয়ে তৈরি। তাই RNA থেকে প্রোটিন পাওয়ার ধাপকে বলে ট্রান্সলেশন বা অনুবাদ। আর এই অনুবাদ করার কাজটি হলো ট্রান্সলেটর বা অনুবাদকের যারা কিনা বিশেষ ধরণের RNA, নাম ট্রান্সফার-RNA বা tRNA। নিউক্লিইক-অ্যাসিড থেকে অ্যামিনো-অ্যাসিডে অনুবাদ করার কাজটি tRNA সম্পাদন করে তার বিশেষ আকৃতির মাধ্যমে। mRNA-এর কোনো বিশেষ আকার নেই, কিন্তু, tRNA অপেক্ষাকৃত ছোট মাত্র ৮০ নিউক্লিয়টাইড লম্বা এবং এর বিশেষ আকার আছে যার মাধ্যমে সে একপ্রান্তে mRNA এবং অন্যপ্রান্তে অ্যামিনো-অ্যাসিডের সাথে যুক্ত হয়। যেকোনো RNA-এর মতোই tRNA তৈরি করা হয় DNA থেকে, কিন্তু, tRNA কখনোই প্রোটিন তৈরির টেম্পলেট (template) বা ছাঁচ হিসেবে ব্যবহৃত হয়না।
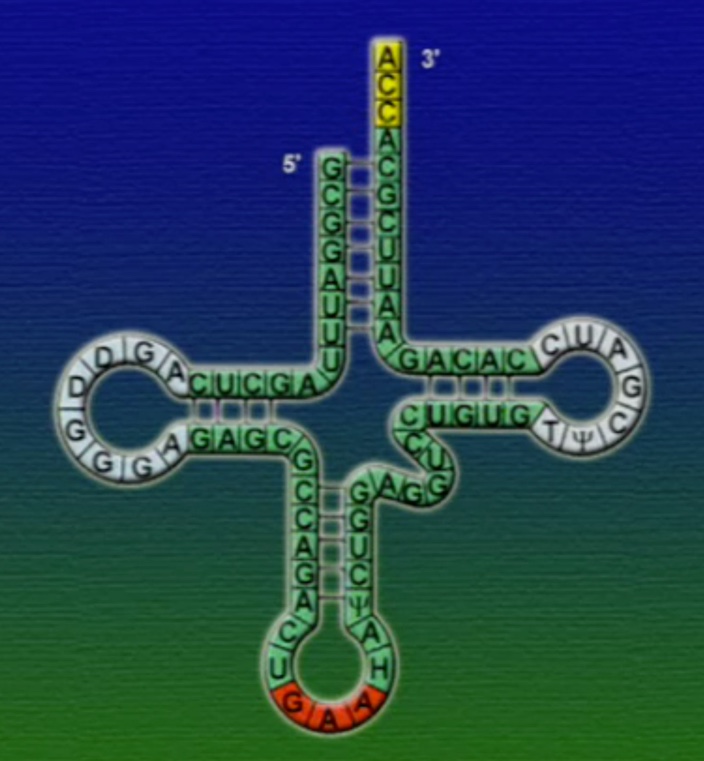
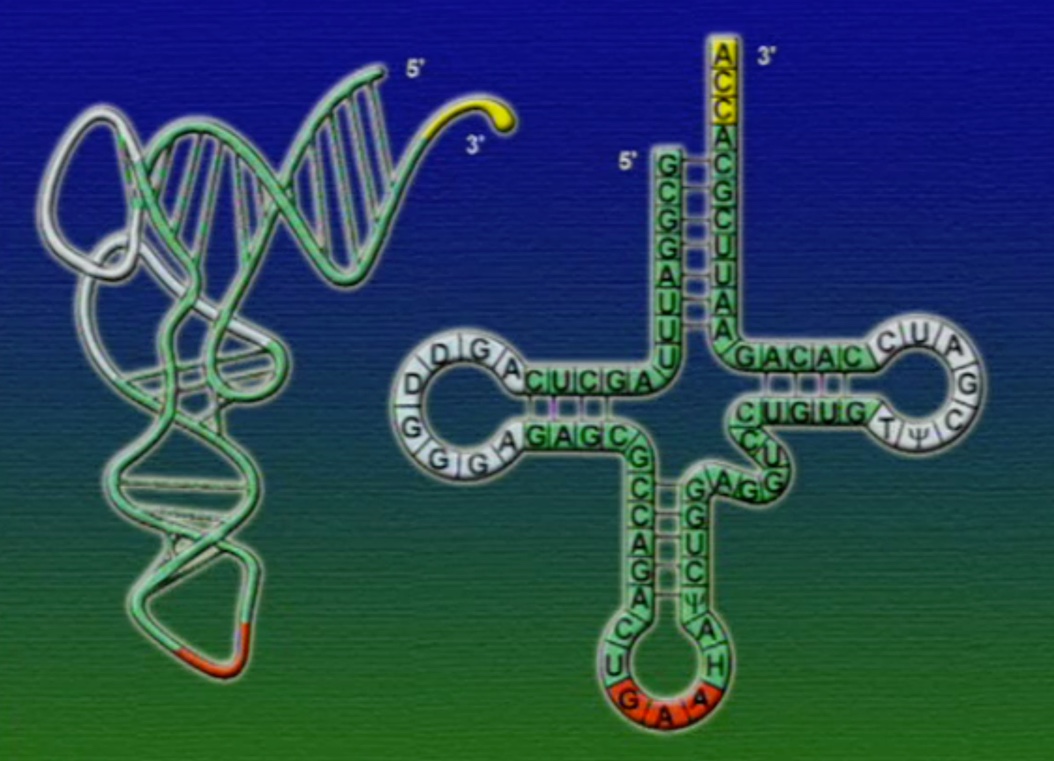
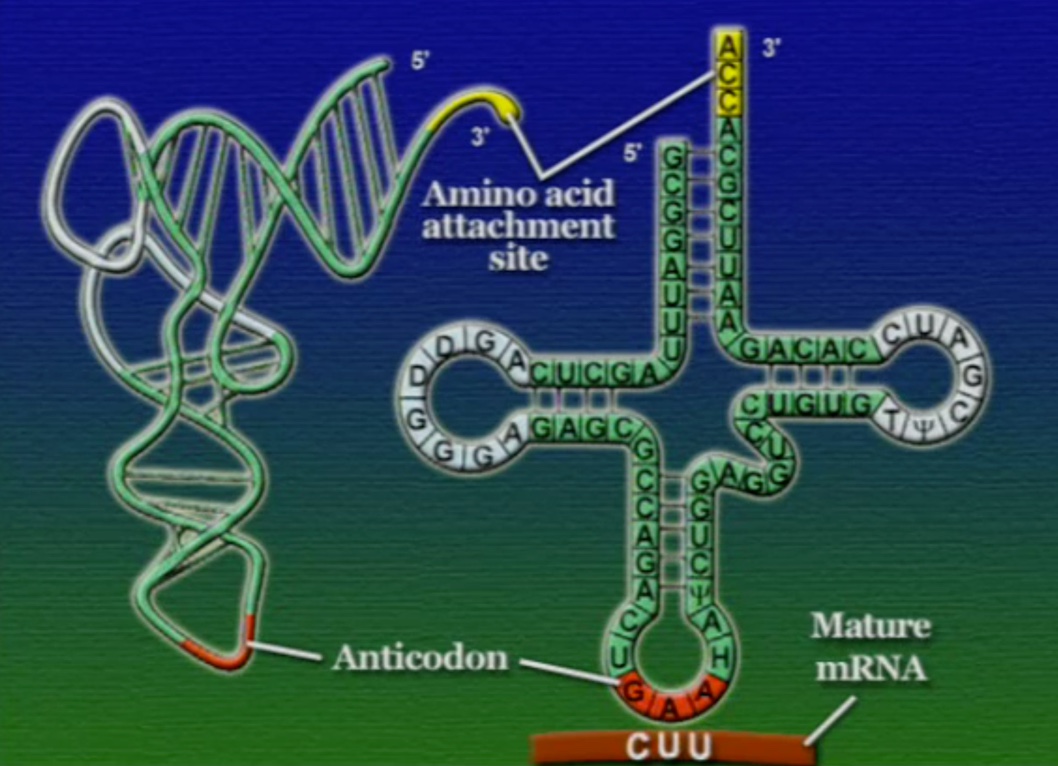
tRNA-এর যে অংশ mRNA-এর কোডোনের সাথে যুক্ত হয় তাকে বলে আন্টিকোডোন (Anticodon), ছবিতে দেখানো GAA অংশটি। মনে হতে পারে, যেহেতু মোট কোডোন সংখ্যা ৬১টি তাই প্রতিটি কোডোনের জন্য আলাদা আলদা tRNA থাকা উচিত। কিন্তু, tRNA মোট ৪৫ ধরণের এবং এদের প্রতিটির আকার একইরকম, পার্থক্য শুধু আন্টিকোডোন অংশটির। আমরা জানি একাধিক কোডোন একটি অ্যামিনো-অ্যাসিডকে সূচিত করতে পারে। এই কোডোনগুলোর পার্থক্য শুধু শেষ বা তৃতীয় নিউক্লিয়টাইডে। অর্থাৎ, কোডোনগুলোর তৃতীয় অবস্থানটি পরিবর্তনশীল। কোডোনগুলোর সাথে তাল মেলাতে tRNA-এর আন্টিকোডোনের শেষ বা তৃতীয় অংশটিও শিথিল (Less critical)। তাই, শুধু একটি অ্যামিনো-অ্যাসিড সূচিত করে এমন অনেকগুলো কোডোনের সাথে tRNA-এর আন্টিকোডোনের অংশটি হাইড্রোজেন বন্ধনে আবদ্ধ হতে পারে। আন্টিকোডোনের তৃতীয় অংশের এই শিথিলতাকে বলে থার্ড-পজিসন-ওয়াবল (Third position wobble)।
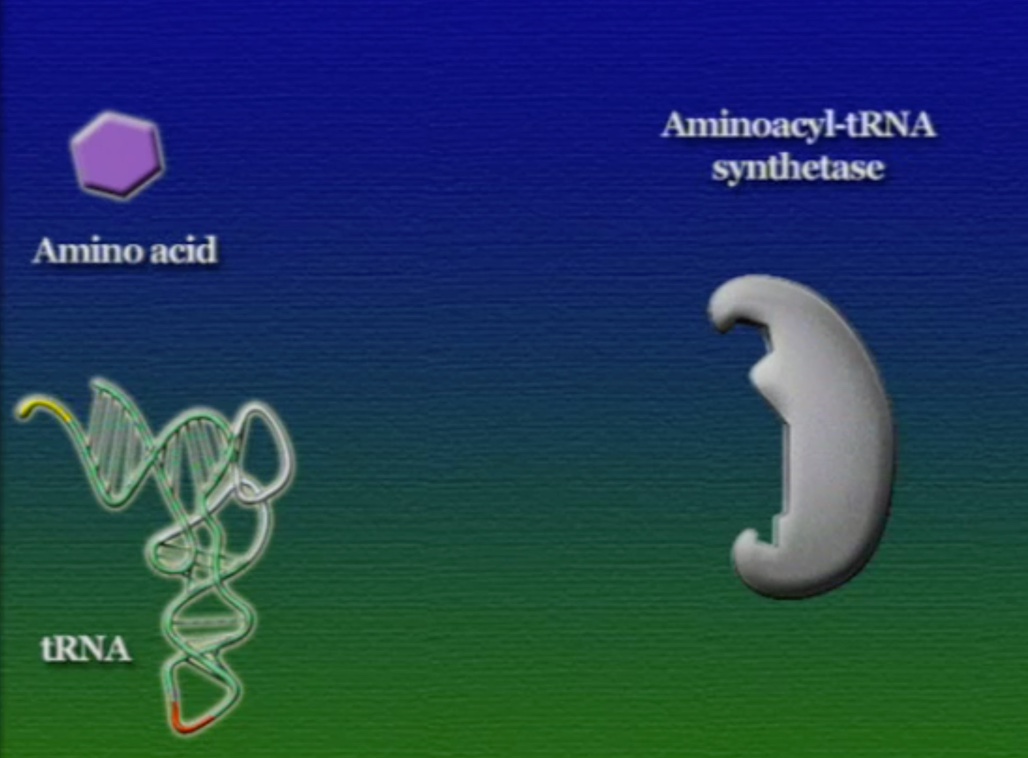
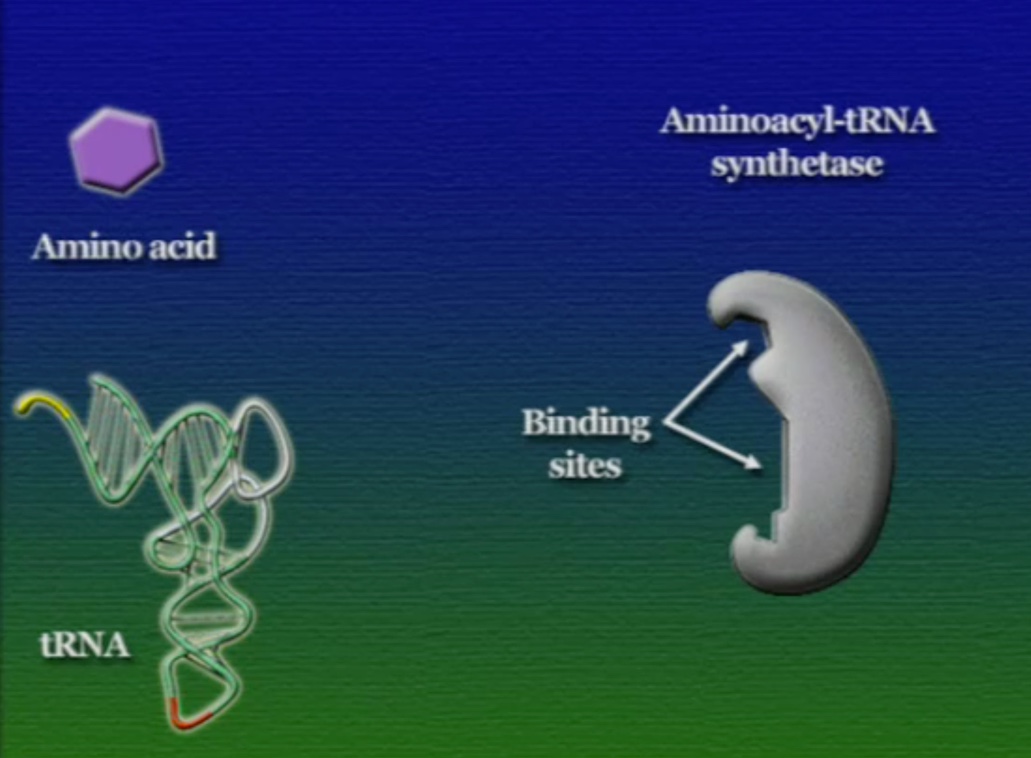
এবার আসা যাক অ্যামিনো-অ্যাসিড সংযুক্তির অংশে, ছবিতে দেখানো ACC অংশটি (Amino acid attachment site)। সমস্যা হলো, প্রত্যেকটি tRNA-এর জন্য এই অংশটি ACC। প্রশ্ন হলো, তাহলে কিভাবে tRNA ২০টি অ্যামিনো-অ্যাসিডের মধ্য থেকে সঠিক অ্যামিনো-অ্যাসিডের সাথে যুক্ত হয়? এই পাজেল সমাধানের দায়িত্ব একটি এনজাইমের নাম অ্যামিনোএসাইল-tRNA-সিস্থেটেজ (Aminoacyl tRNA synthetase)। এই এনজাইমটি অনেকটা বিয়ের ঘটকের মতো। অ্যামিনোএসাইল-tRNA-সিস্থেটেজের আকৃতিতে দুটো সংযুক্তিস্থল থাকে, একটা অংশে একটা নির্দিষ্ট tRNA যুক্ত হয়, অন্য অংশে যুক্ত হয় একটা নির্দিষ্ট অ্যামিনো-অ্যাসিড। প্রতিটি tRNA-এর জন্য আলাদা আলাদা অ্যামিনোএসাইল-tRNA-সিস্থেটেজ এনজাইম রয়েছে। অ্যামিনোএসাইল-tRNA-সিস্থেটেজের মাধ্যমে tRNA এবং অ্যামিনো-অ্যাসিডের বন্ধন তৈরি প্রক্রিয়াটি নিচের ছবিগুলোর মাধ্যমে দেখানো হলো।
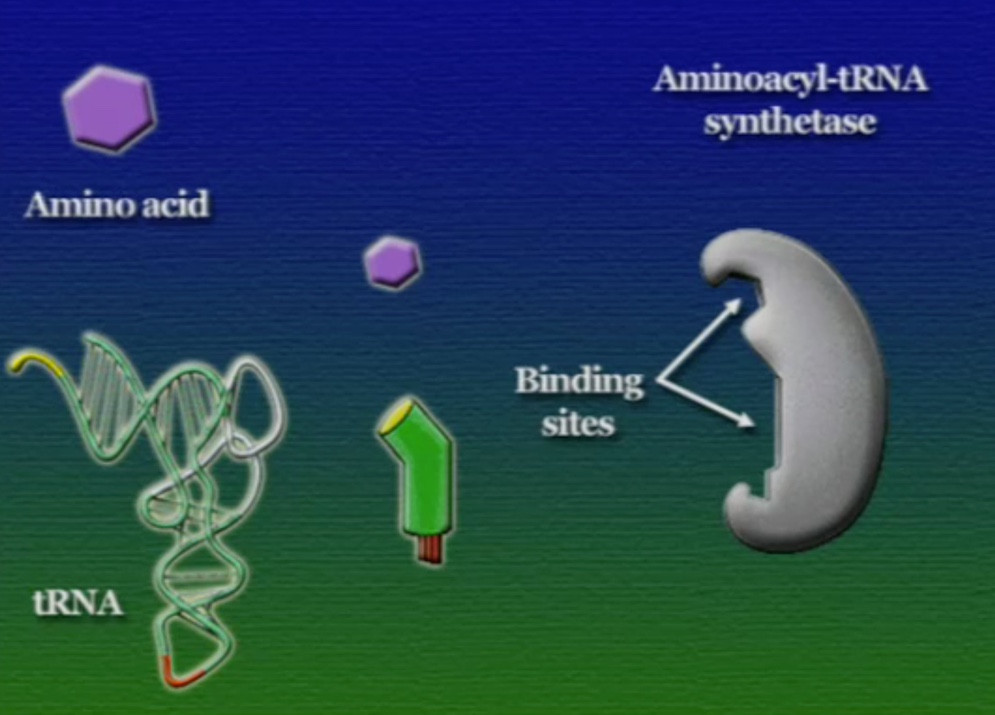
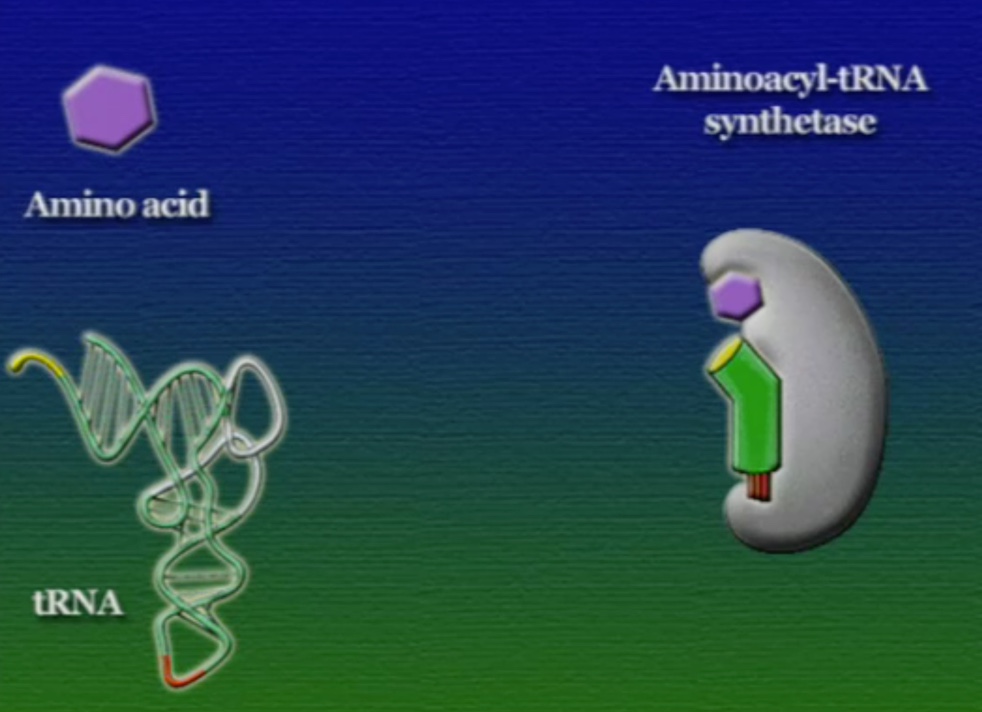
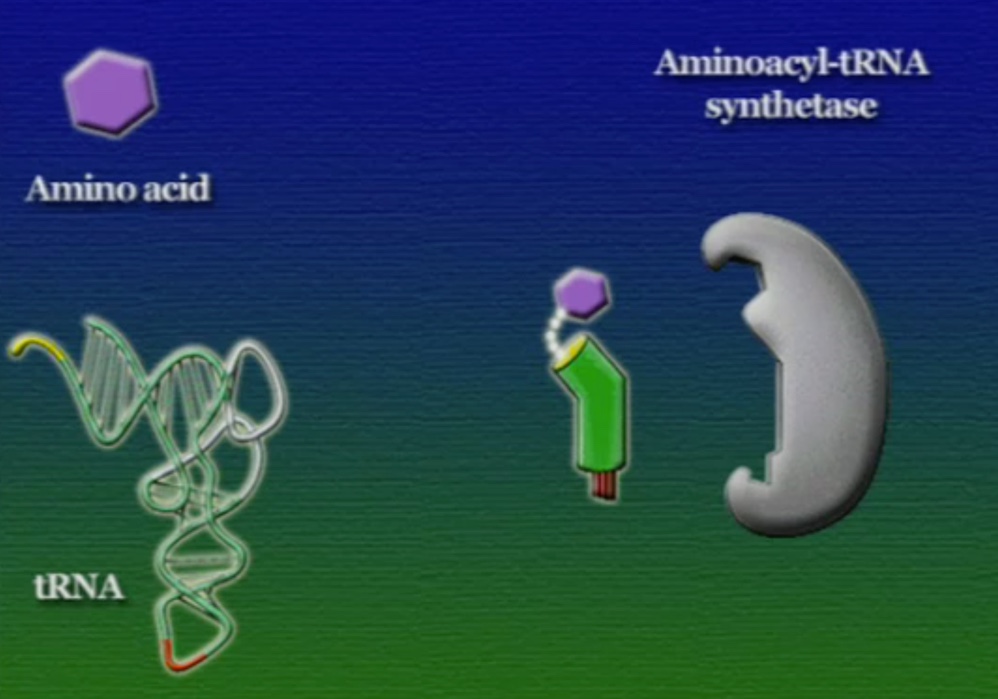
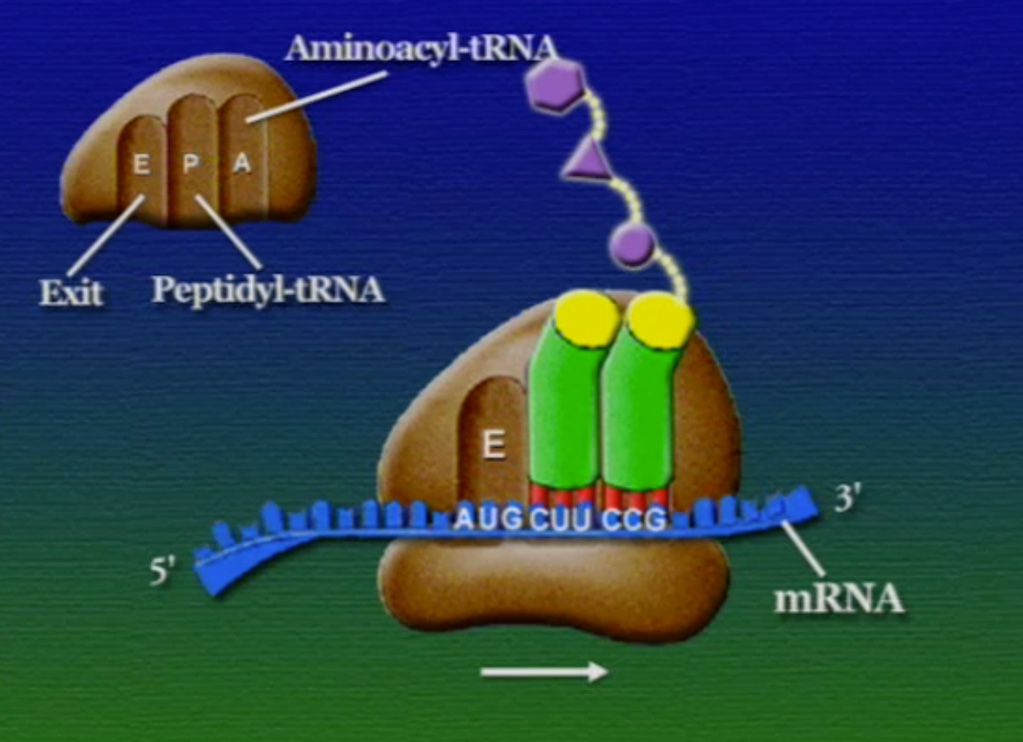
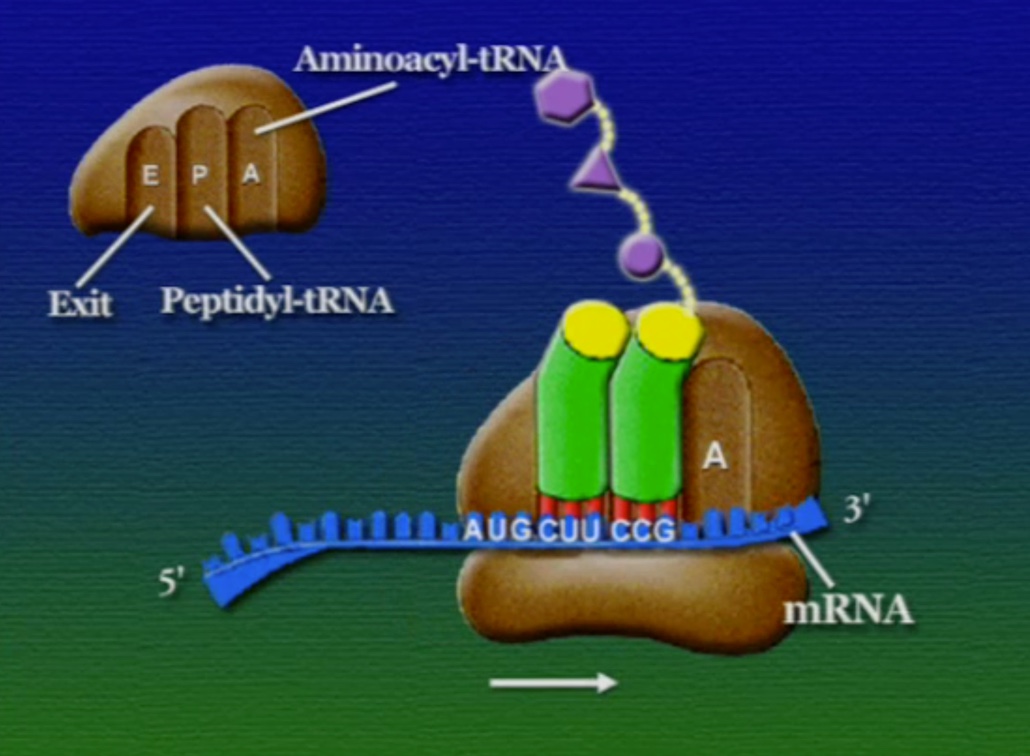
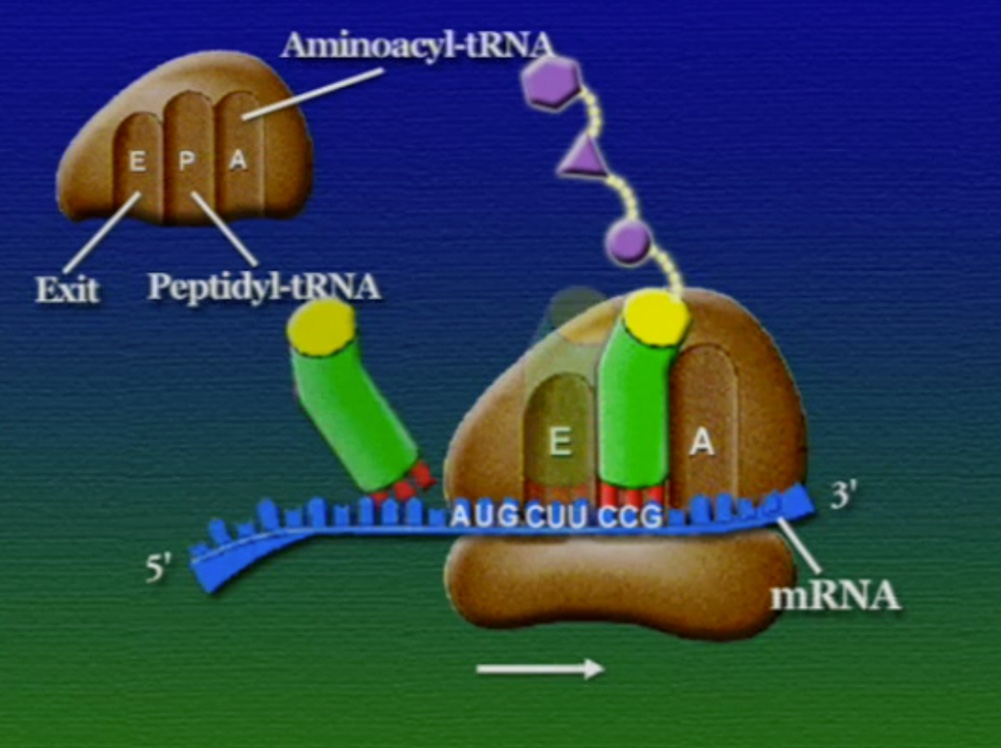
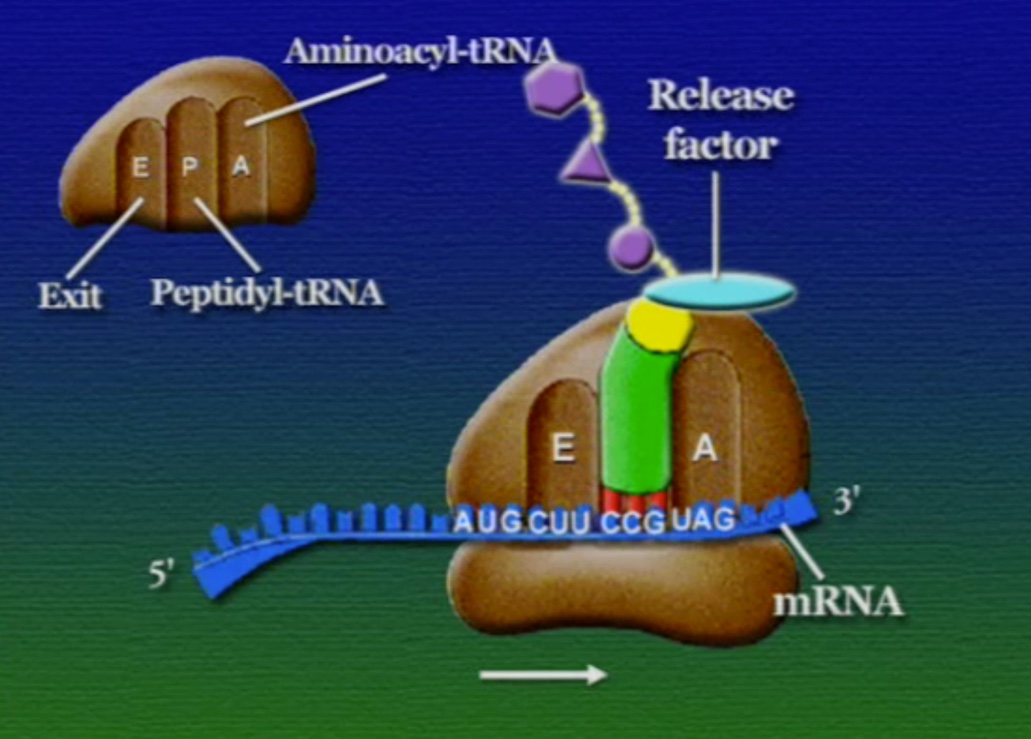
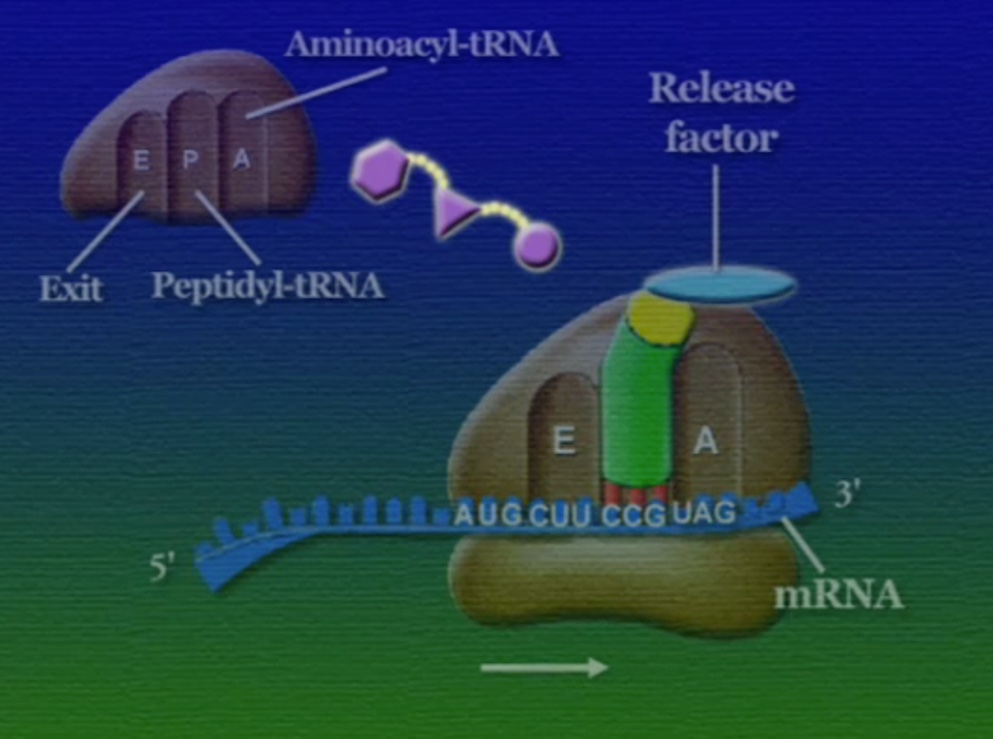
অ্যামিনো-অ্যাসিডের সাথে যুক্ত হবার পর এই tRNA-দের বলে অ্যামিনোএসাইল-tRNA এবং এদের সরাসরি পাঠানো হয় রাইবোসোমে। রাইবোসোম ৬০% তৈরি রাইবোসোমাল-RNA (Ribosomal RNA) বা rRNA এবং ৪০% বিভিন্ন এনজাইম দিয়ে তৈরি। রাইবোসোম হলো প্রোটিন তৈরির যন্ত্র, এরা অত্যন্ত দ্রুত কাজ করে। এদের দ্রুততার কারণেই একটি কোষ প্রতি সেকেন্ডে প্রায় দেড় লক্ষ্য প্রোটিন তৈরি করতে পারে। রাইবোসোমের দুটি অংশ আছে। বড় অংশটির তিনটি প্রক্রিয়াজাতকরণ অঞ্চল আছে, যাদের নাম ছবি অনুসারে, A = অ্যামিনোএসাইল-tRNA, P = পেপটাইডিইল-tRNA, E = এক্সিট।
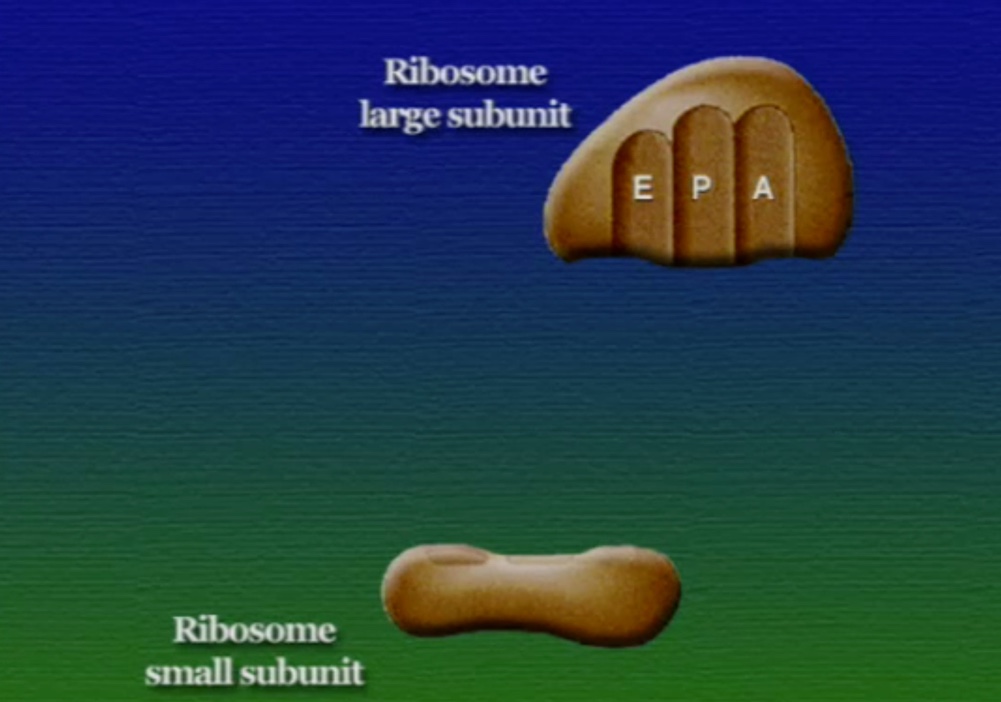
রাইবোসোমে প্রোটিন তৈরি হয় তিনটি ধাপে। শুরু (Initiation), দীর্ঘায়িতকরণ (Elongation), এবং সমাপ্তি (Termination)। নিচে ধাপগুলো ছবির মাধ্যমে দেখানো হলো।
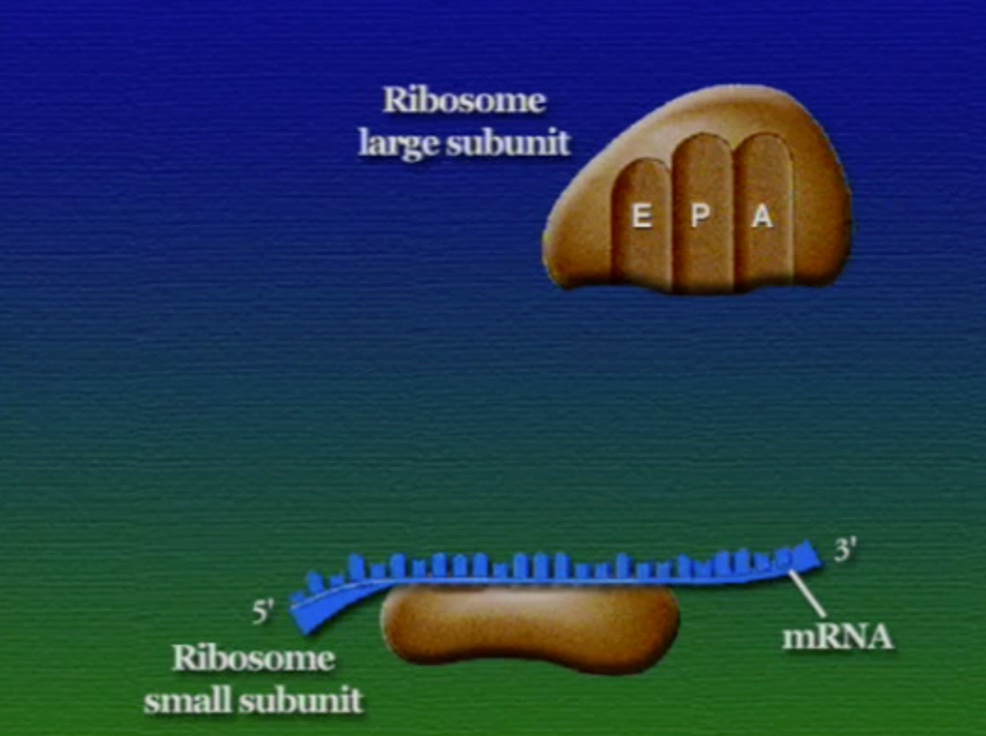
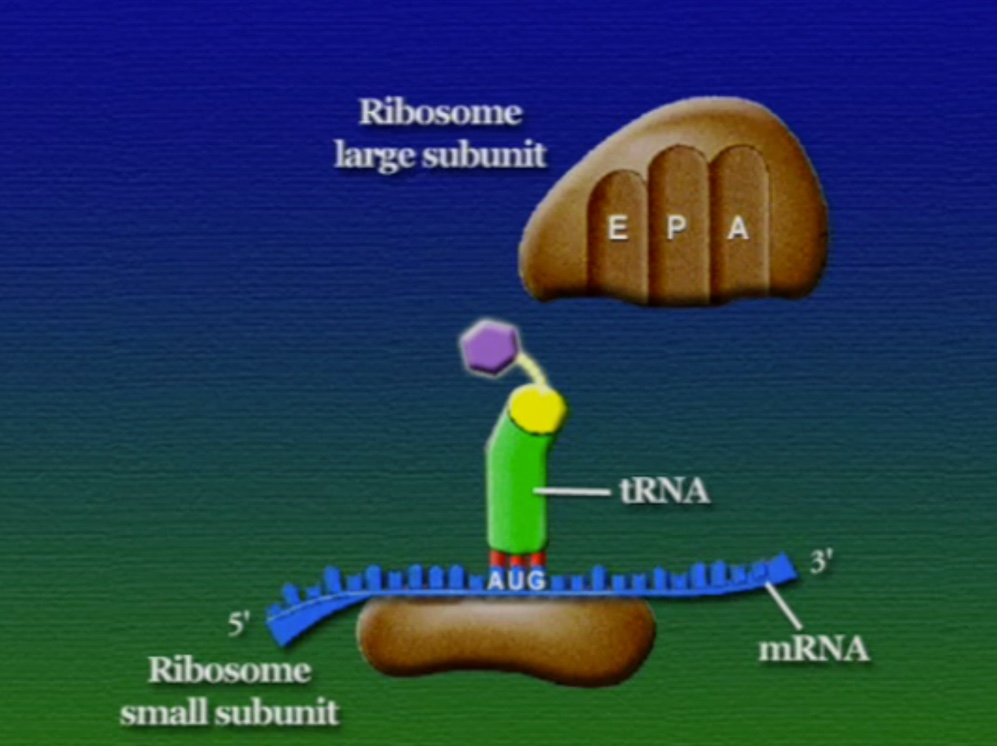
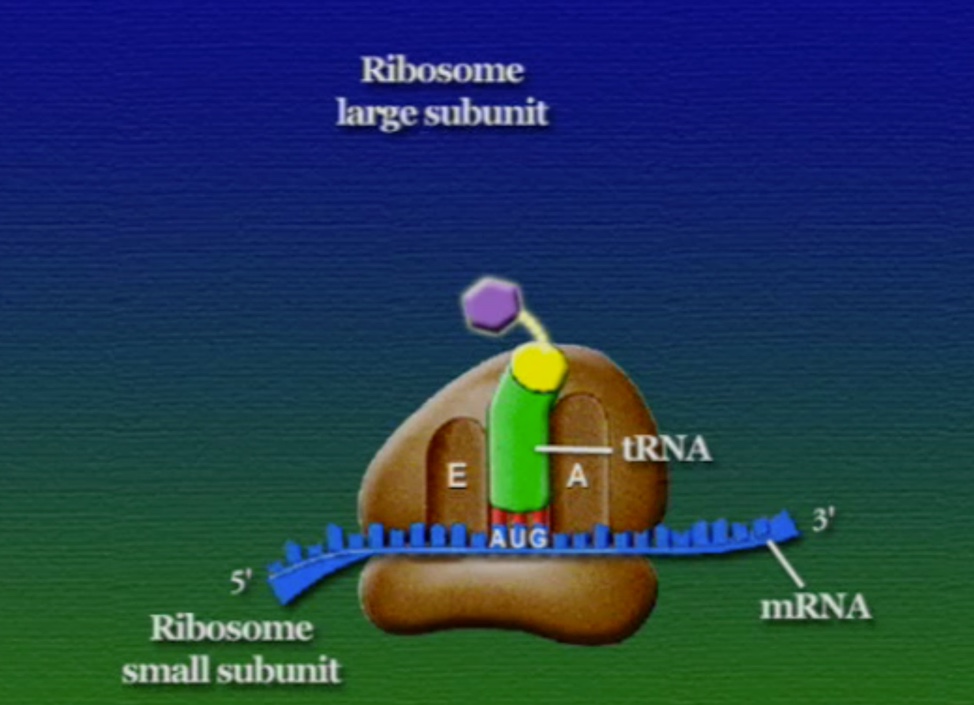
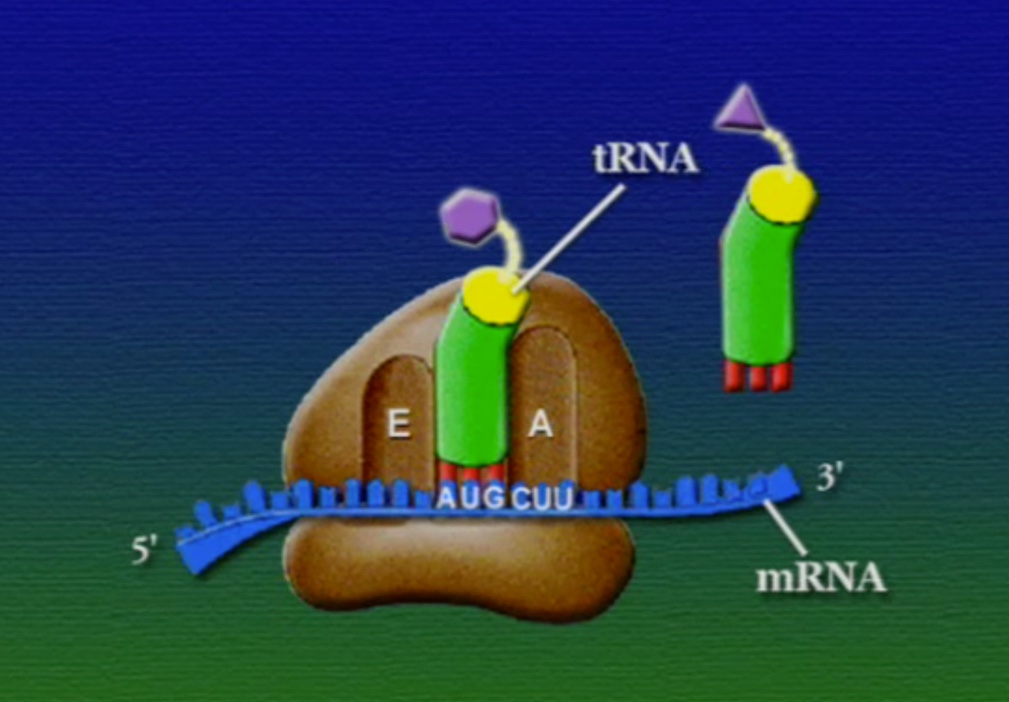
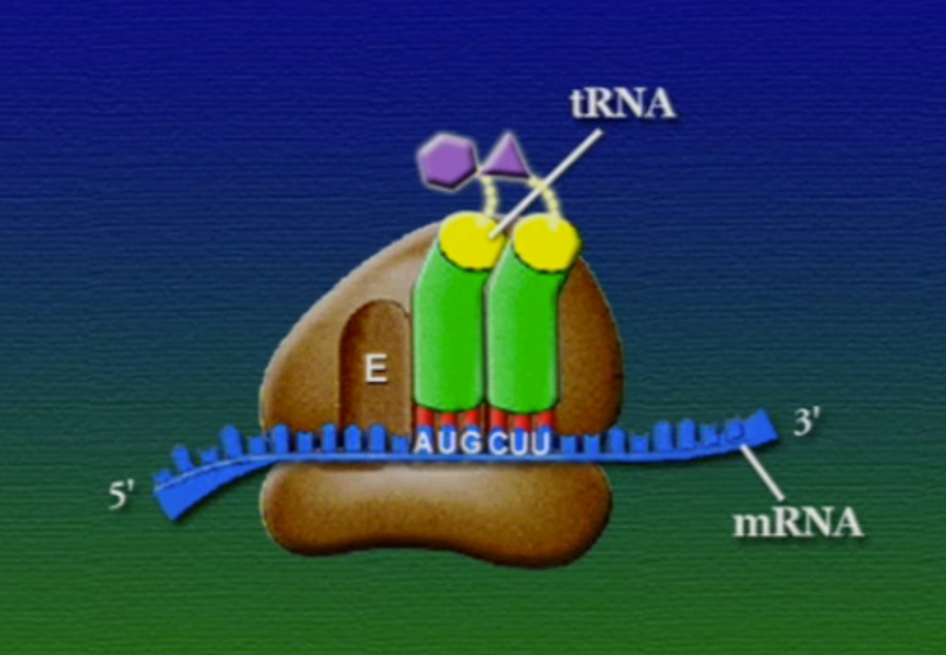
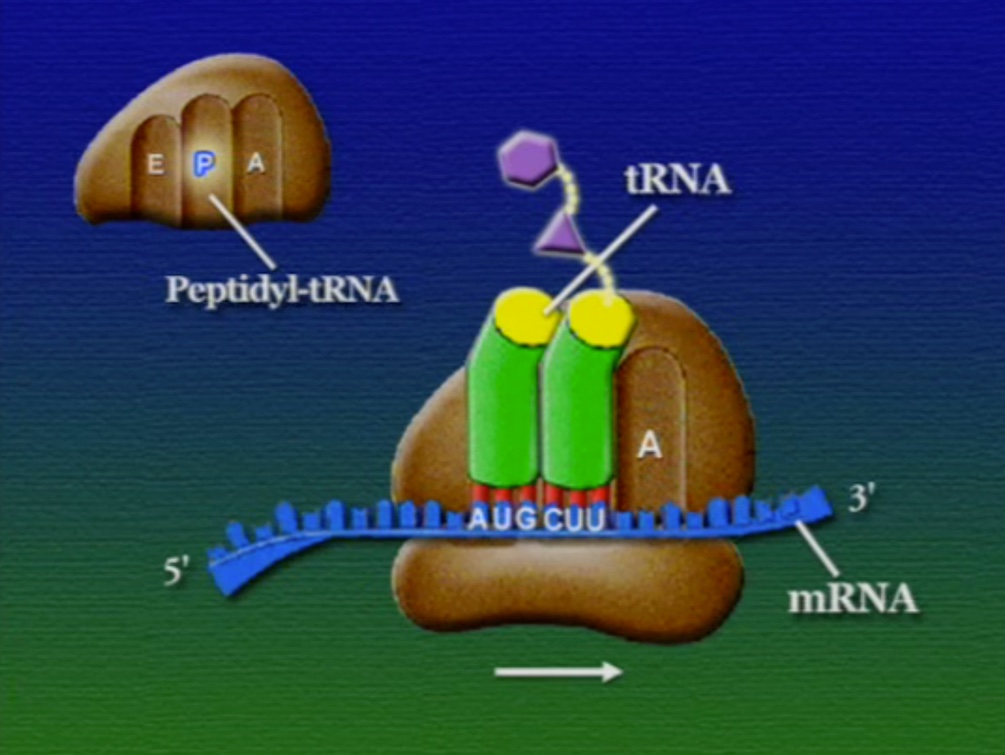
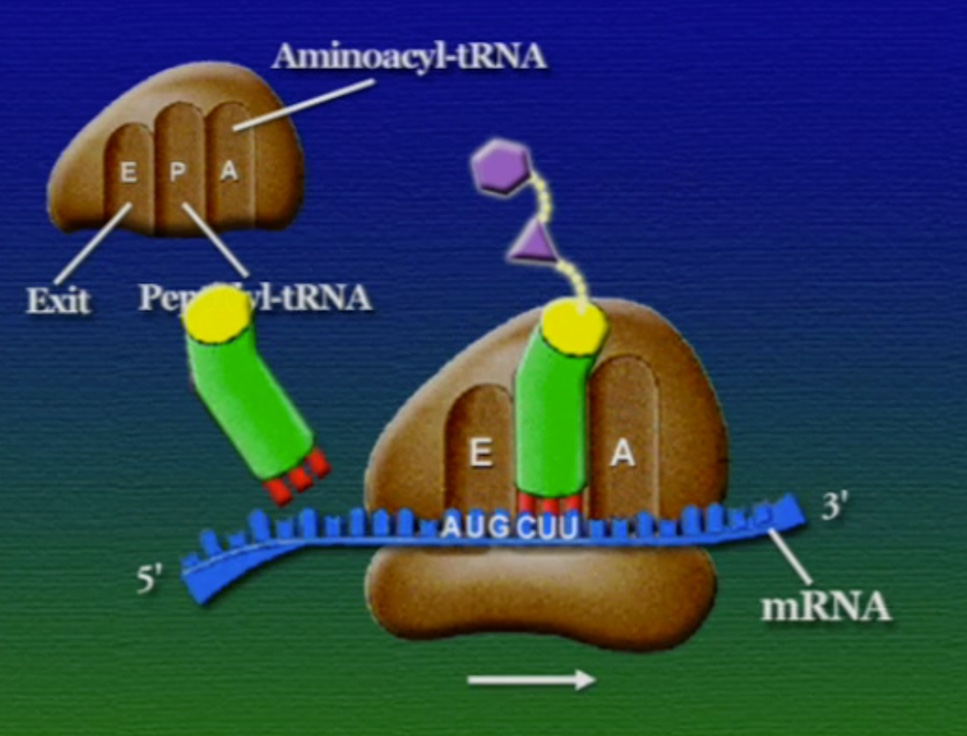
লম্বা হতে থাকা পেপটাইড-চেইনটি সবসময় P অঞ্চলে যুক্ত থাকে তাই একে পেপটাইডিইল-tRNA বলে। A অঞ্চলে যুক্ত হয় নতুন নতুন অ্যামিনোএসাইল-tRNAগুলো। আর অ্যামিনো-অ্যাসিড হারানো tRNAগুলো E অঞ্চলে এসে বেরিয়ে যায়, তাই একে এক্সিট বলে।
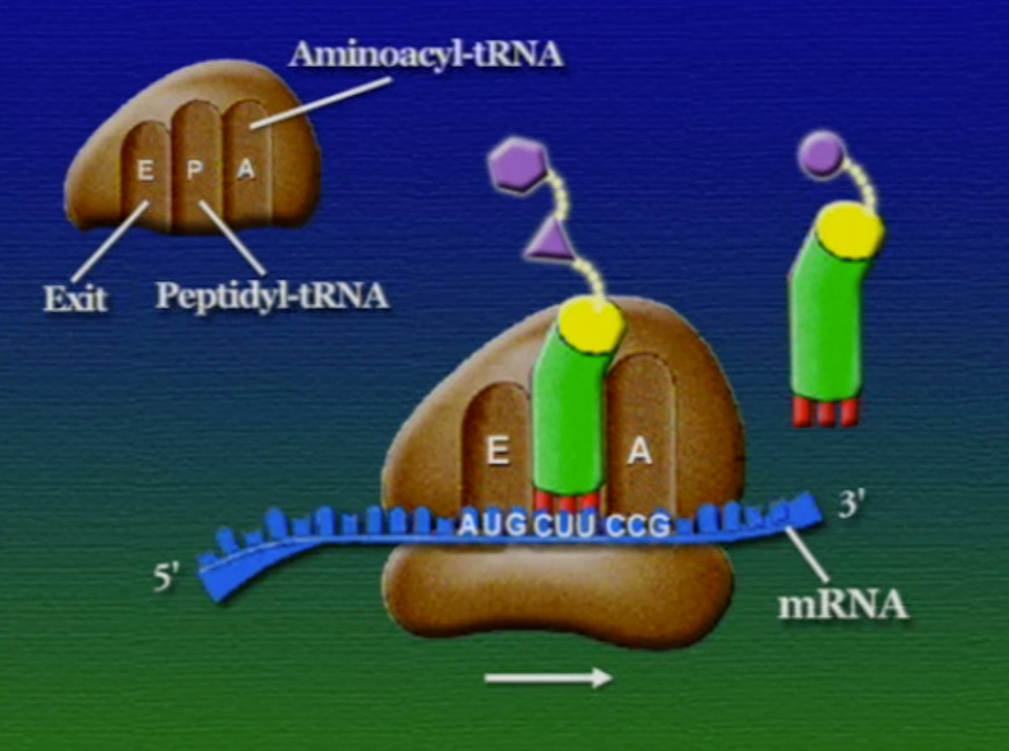
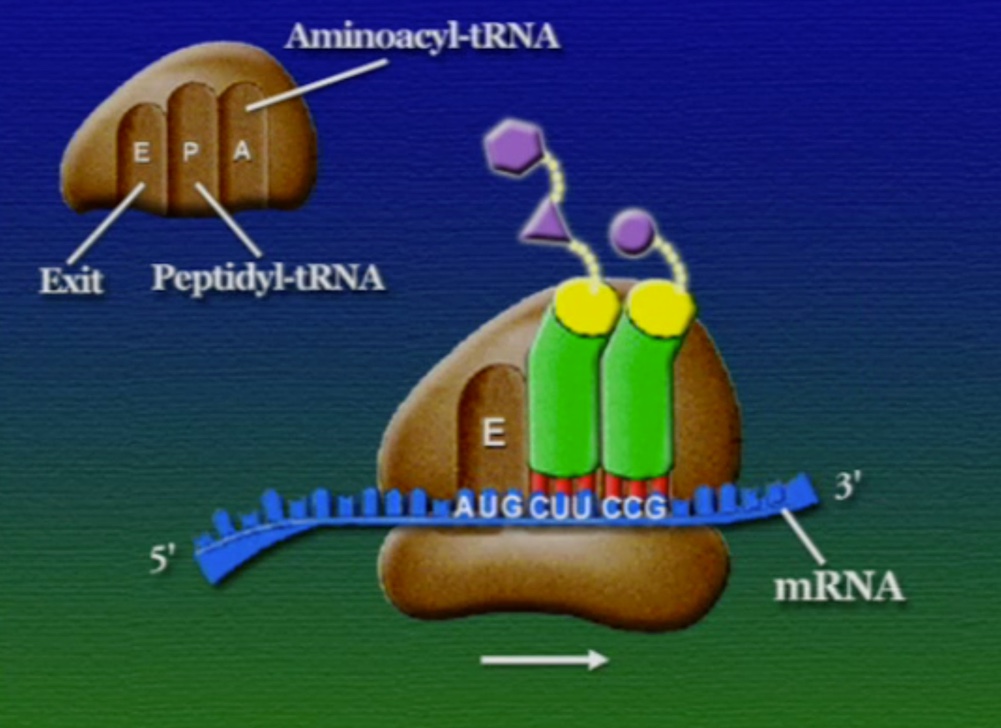
প্রোটিনের পেপটাইড-চেইন তৈরি হয়ে গেলে, এদের পাঠানো হয় কোষের বিশেষ একটা অংশে যেখানে এদের প্যাকেটজাত করা হয়। আগে বলা হয়েছিলো, প্রোটিনের কার্যকারিতা নির্ভর করে তার আকারের ওপর। বিশেষ করে তাদের টারশিয়ারী স্ট্রাকচার খুব গুরুত্বপূর্ণ। সমস্যা হলো, পেপটাইড-চেইন নিজে থেকেই ঠিক মতো গুটিয়ে যেতে পারেনা। পেপটাইড-চেইনকে ঠিক মতো ভাঁজ করার জন্য দরকার একটি সাহায্যকারী প্রোটিনে যার নাম স্যাপারন প্রোটিন (Chaperone protein)। আরেকটা ব্যাপার, প্রতিটি পেপটাইড চেইনের শুরু মিথাইয়ােনিন দিয়ে (কারণ স্টার্ট-কোডোন AUG মিথাইয়ােনিন অ্যামিনো-অ্যাসিড সূচিত করে), কিন্তু, অনেক প্রোটিনের এই শুরুর মিথাইয়ােনিন দরকার হয় না। সুতরাং, এই প্যাকেটজাত প্রক্রিয়ায় কিছু কিছু ক্ষেত্র এই মিথাইয়ােনিন কেটে বাদ দেয়া হয়। কোষের এই প্যাকেটজাতকরণ অংশটিকে বলে এন্ডোপ্লাসমিক-রেটিকাম (Endoplasmic reticulum)। আমরা যে রাইবোসোমের কথা বলছিলাম, তারা মূলত এন্ডোপ্লাসমিক-রেটিকামের সাথে যুক্ত থাকে। কিছু কিছু রাইবোসোম সাইটোপ্লাসোমে ভেসে বেড়ায়, তারা যে প্রোটিন তৈরি করে সেগুলোর বিশেষ কোনো পোস্ট-প্রসেসিং-এর মধ্য দিয়ে যেতে হয় না। এন্ডোপ্লাসমিক-রেটিকামের রাইবোসোমযুক্ত অংশকে বলে রাফ-এন্ডোপ্লাসমিক-রেটিকাম (Rough -endoplasmic reticulum) বা RER। রাইবোসোম থেকে মুক্ত হয়ে পেপটাইড-চেইন এই এন্ডোপ্লাসমিক-রেটিকামে প্রবেশ করে এবং পরবর্তী প্রক্রিয়াজাতকরণ ধাপগুলো চলতে থাকে।
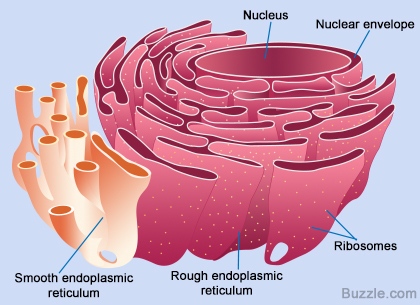
উৎপাদিত কিছু প্রোটিন কোষের ভেতরেই কাজে লাগে আর কিছু প্রোটিন নিউক্লিয়াসে, এমনকি কোষের বাহিরেও পাঠাতে হয়। তাই, প্রোটিনকে তার সঠিক গন্তব্যে পৌঁছানোর জন্য প্রোটিনের গায়ে তার গন্তব্যের ঠিকানা লাগিয়ে দিতে হয়। এন্ডোপ্লাসমিক-রেটিকামকে একটা প্যাকেজিং কারখানার সাথে তুলোনা করা চলে। এর ভেতরের আঁকাবাঁকা পথগুলো যেনো কারখানার একেকটা ওয়ার্কস্টেশন, কোনো অংশে পেপটাইড-চেইনকে ভাঁজ করা হয়, কোনো অংশে কাটছাট করা হয়, আবার কোনো অংশে গ্লুকোসের প্রলেপ লাগানো হয় ইত্যাদি ইত্যাদি।
ক্রিকের সেন্ট্রাল ডগমার একটা উল্লখযোগ্য দিক হলো, এটা ল্যামার্কিয়ান বিবর্তনকে ভুল প্রমাণিত করে। ল্যামার্কিয়ান বিবর্তন অনুসারে, পরিবেশগত অভিযোজন বংশপরম্পরায় পরিচালিত হয়। কিন্তু, সেন্ট্রাল ডগমা অনুসারে, একবার প্রোটিন তৈরি করা হলে, সেই প্রোটিনের তথ্য কখনোই DNA-তে সংযুক্ত করা যায় না।
অবলম্বনে: “The Great Courses” থেকে প্রকাশিত “Biology: The Science of Life” by “Stephen Nowicki”