আমাদের জন্ম একটা নিষিক্ত ডিম্বাণু থেকে, যাকে বলে জাইগোট (Zygote)। এই জাইগোট বিভাজনের মাধ্যমে আমরা একটা বহুকোষী আকৃতি পাই, যেখানে থাকে হৃৎপিন্ড, যকৃত, কিডনি, চোখ ইত্যাদির মতো বিশেষায়িত অঙ্গগুলো। এই অঙ্গগুলোর মূল পার্থক্য এদের উৎপাদিত এবং ব্যবহৃত প্রোটিনে। আর এই প্রোটিনগুলো তৈরি হয় নির্দিষ্ট জিন থেকে। জাইগোট বিভাজনে প্রতিটি বিভাজিত কোষ তার মাতৃকোষের হুবুহু নকল DNA-কপি পায়। ফলে, আমাদের প্রতিটি দেহকোষের DNA হুবুহু এক। ব্যতিক্রম শুধু শুক্রাণু এবং ডিম্বাণুর ক্ষেত্রে। প্রশ্ন হলো, কিভাবে কোনো নির্দিষ্ট অঙ্গ শুধুই প্রয়োজনীয় প্রোটিন উৎপাদন করে এবং অপ্রয়োজনীয় প্রোটিন উৎপাদন থেকে বিরত থাকে? প্রোটিন উৎপাদন বিষয়টি নিয়ন্ত্রিত হয় কোষ পর্যায়ে, প্রতিটি কোষ কঠিনভাবে ট্রান্সক্রিপশন ধাপ নিয়ন্ত্রণের মাধ্যমে প্রয়োজনীয় প্রোটিন উৎপাদন নিয়ন্ত্রণ করে।
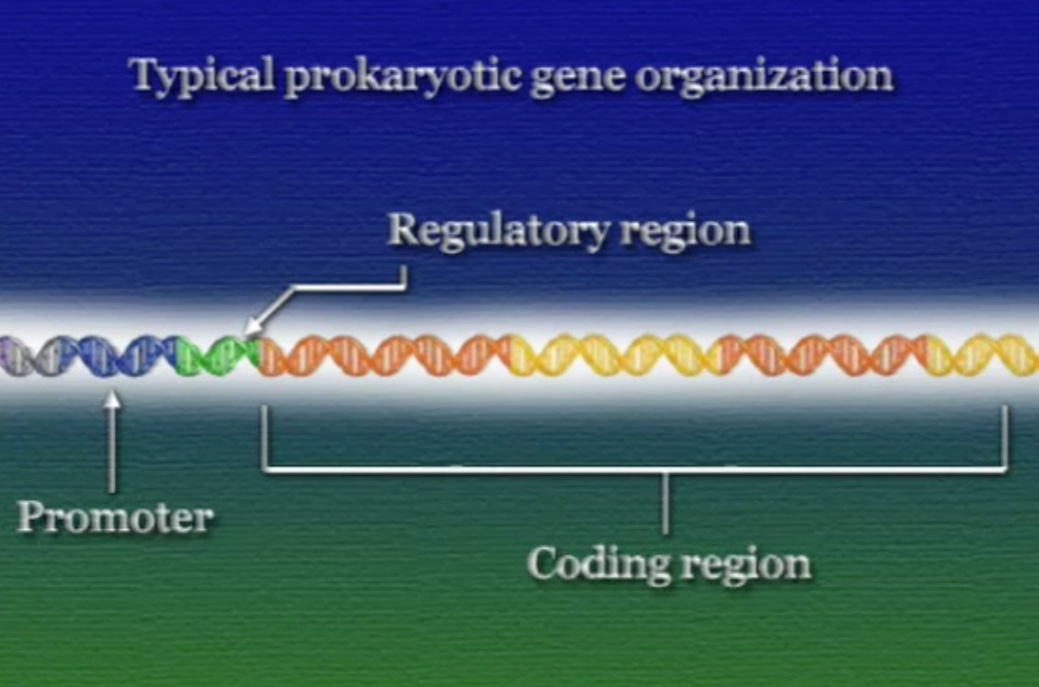
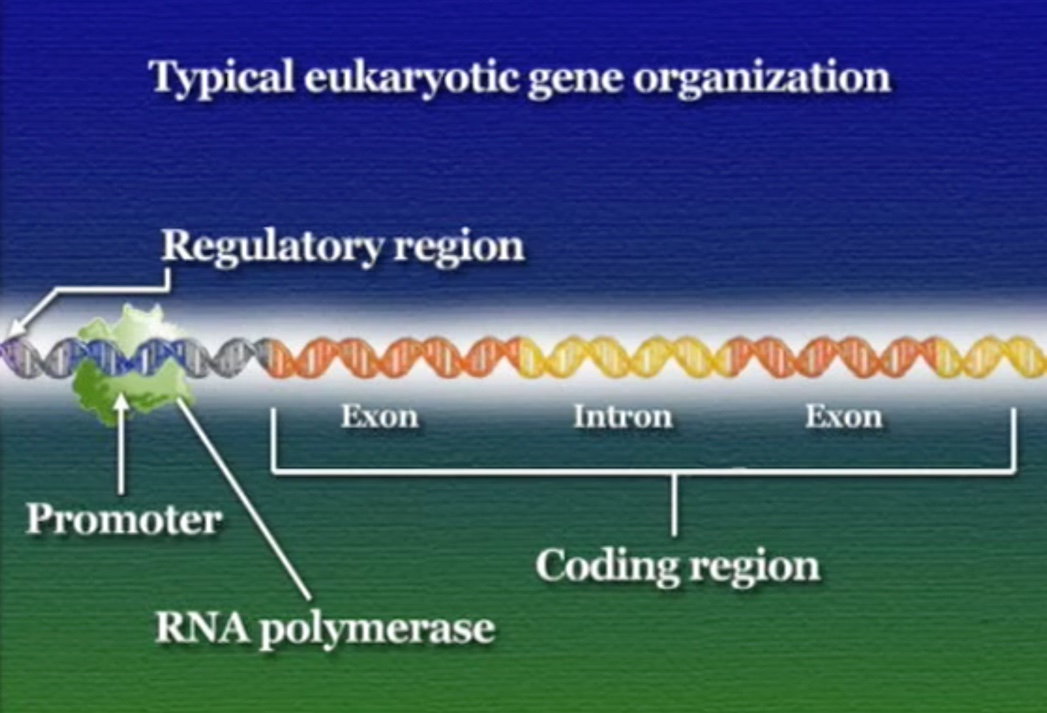
ট্রান্সক্রিপশন নিয়ন্ত্রণের জন্য প্রতি জিনের এক বা একাধিক নিয়ন্ত্রক-অঞ্চল বা রেগুলেটরি-রিজিয়ন (Regulatory region) থাকে। এই রেগুলেটরি-রিজিয়নগুলো কোডিং-রিজিয়নের সম্মুখে (Upstream) অবস্থিত। রেগুলেটরি-রিজিয়নের মাধ্যমে ট্রান্সক্রিপশন নিয়ন্ত্রণ করার উপায়গুলো প্রোক্যারিয়টিক ও ইউক্যারিয়টিক কোষ ভেদে ভিন্ন ভিন্ন হয়।
আমরা জানি, ট্রান্সক্রিপ্ট ধাপে RNA-পলিমাররেস জিনের প্রমোটর অঞ্চলে যুক্ত হয় এবং কোডিং-রিজিয়ন ধরে সামনে এগুতে থাকে। প্রোক্যারিয়টদের জিনে রেগুলেটরি-রিজিয়নটি প্রমোটর ও কোডিং-রিজিয়ন দুটির মাঝে অবস্হিত।
প্রোক্যারিয়টেরা যখন কোনো জিনের ট্রান্সক্রিপশন বন্ধ করতে চায়, তখন তারা একটি বিশেষ ধরণের প্রোটিন তৈরি করে যেটা জিনের রেগুলেটরি-রিজিয়ন অঞ্চলে আটকে থাকে। আমরা জানি, প্রতিটি প্রোটিনের একটি স্বাতন্ত্র্য (Unique) আকার আছে। অন্যদিকে জিনের রেগুলেটরি-রিজিয়নগুলো DNA-ডাবল-হেলিক্সের অংশ, ফলে প্রতিটি রেগুলেটরি-রিজিয়নেরও একটি নির্দিষ্ট কাঠামো আছে। প্রোটিনের সাথে DNA-এর এই বন্ধন অত্যন্ত সুনির্দিষ্ট (Very specific), অর্থাৎ, চাইলেই যেকোনো প্রোটিন যেকোনো রেগুলেটরি-রিজিয়নে আটকাতে পারেনা। এই প্রোটিনদের বলে রেগুলেটরি-প্রোটিন (Regulatory protein)। যখন এই রেগুলেটরি-প্রোটিন রেগুলেটরি-রিজিয়নে আটকে থাকে তখন তারা শারীরিকভাবে RNA-পলিমারেসকে সামনে এগুতে বাধা দেয়। এই ধরণের জিন নিয়ন্ত্রণকে বলে নেগেটিভ-কন্ট্রোল (Negative control)।
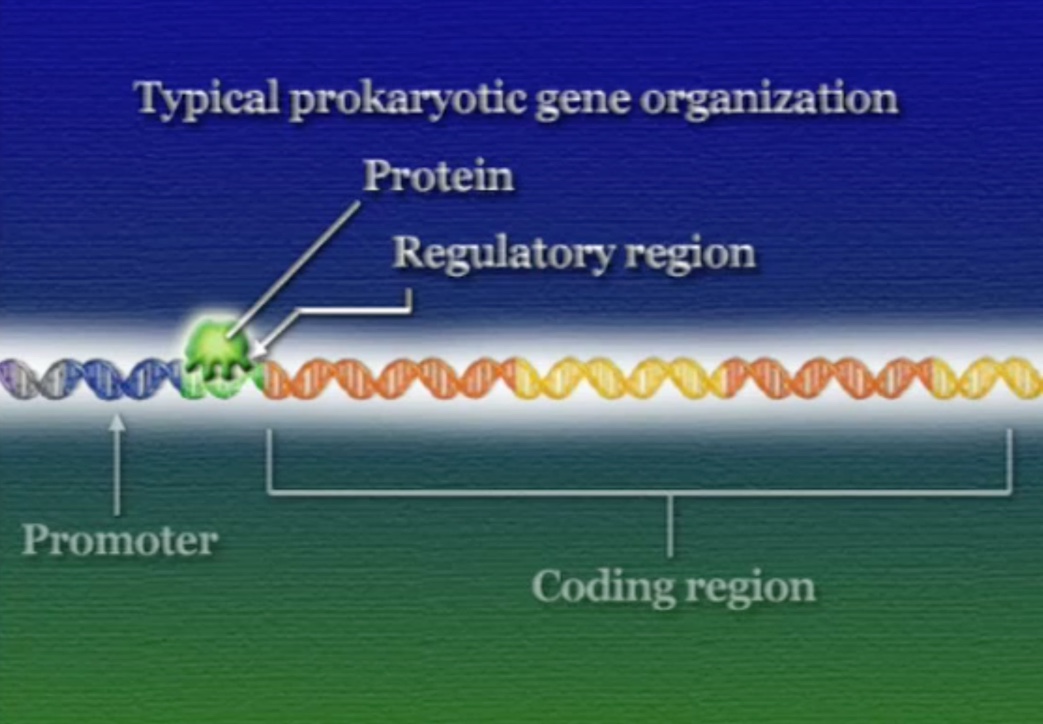
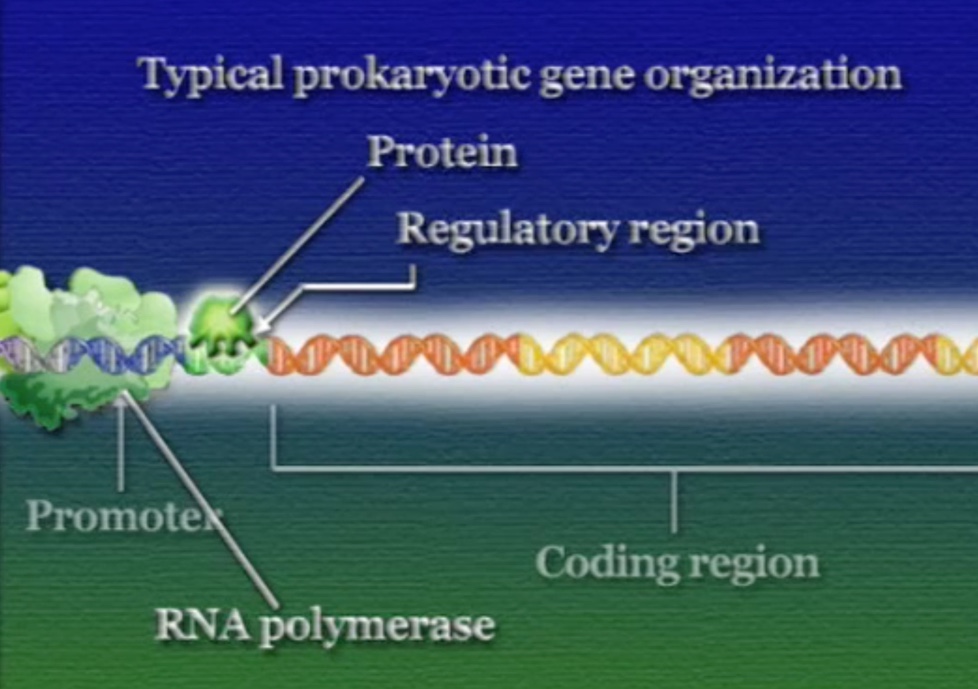
ইউক্যারিয়টদের পদ্ধতি ঠিক উল্টো। ইউক্যারিয়টিক কোষে রেগুলেটরি-প্রোটিন আসলে RNA-পলিমারেসকে প্রমোটর খুঁজে পেতে সাহায্য করে। রেগুলেটরি-রিজিয়নে লেগে থাকা একাধিক রেগুলেটরি-প্রোটিনের ওপর নির্ভর করে RNA-পলিমারেসের প্রমোটর খুঁজে পাওয়া। মনে রাখা দরকার, RNA-পলিমারেসের কোনো বাছবিচার নেই, সে যেই জিনের প্রমোটরের সাথে আটকাতে পারবে RNA-পলিমারেস সেই জিনকেই ট্রান্সক্রিপ্ট করা শুরু করবে। ইউক্যারিয়টদের এই নিয়ন্ত্রণ পদ্ধতিকে বলে পজিটিভ-কন্ট্রোল (Positive control)।
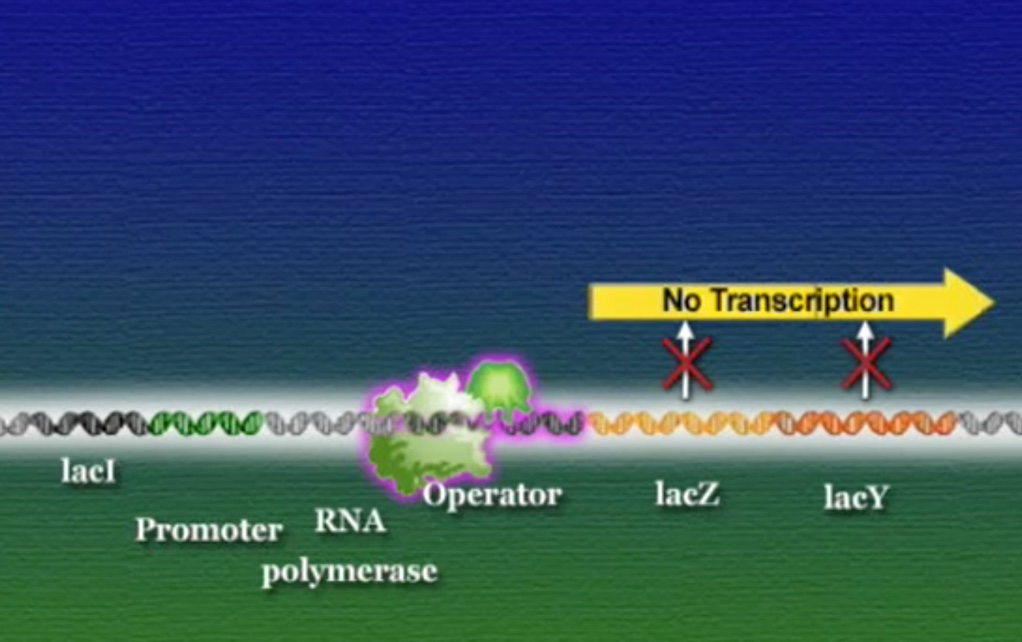
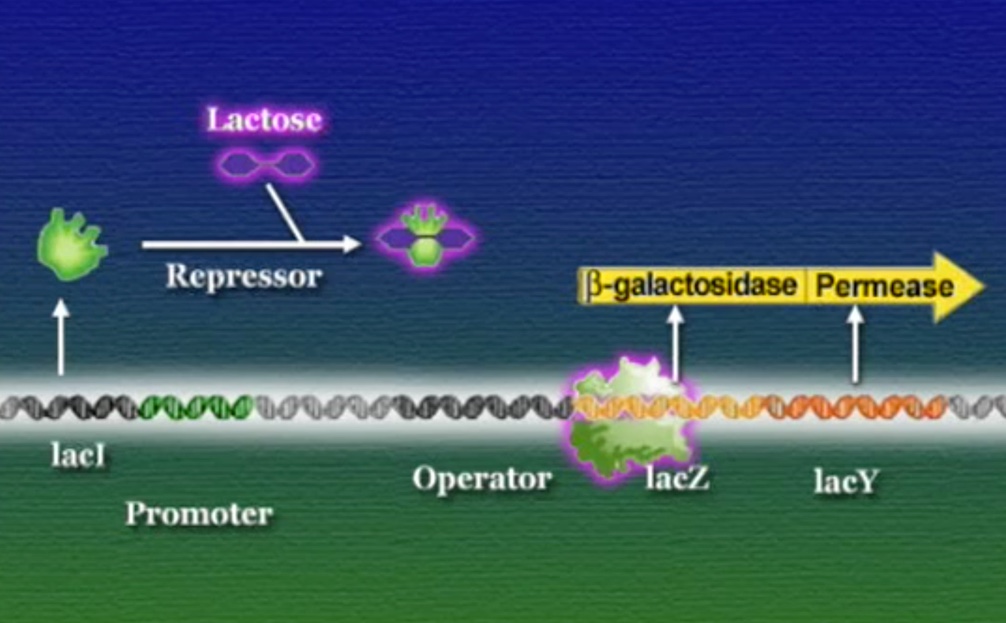
এবার আসা যাক একটা বাস্তব উদাহরণে, ই-কোলাই আমাদের অন্ত্রে বসবাসকারী ব্যাকটেরিয়া। এরা প্রোক্যারিয়ট, মূলত গ্লুকোস খাবার হিসেবে গ্রহণ করে। কিন্তু, আমরা যখন দুধ বা দুগ্ধজাত খাবার খাই তখন এরা দুধের ল্যাক্টোসকেও খাবার হিসেবে গ্রহণ করতে পারে। ল্যাক্টোস (Lactose) একটা জটিল সুগার। ই-কোলাই প্রথমেই ল্যাক্টোসকে ভেঙ্গে দুটি সরল সুগারের পরিণত করে, গ্লুকোস ও গ্যালাকটোসে (Galactose)। এই ল্যাক্টোস ভাঙ্গার জন্য ই-কোলাই যে এনজাইম তৈরি করে তার নাম বেটা-গ্যালাকটোসাইডেস (β-galactosidase)। পাশাপাশি ই-কোলাই আরেকটি এনজাইম ব্যবহার করে, গ্যালাকটোসাইড-পারমিয়েস (Galactoside permease) যার কাজ ল্যাক্টোসকে কোষের বাহির থেকে ভেতরে আনা। মজার ব্যাপার হলো, ই-কোলাই তখনই এই এনজাইম দুটি তৈরি করে যখন ল্যাক্টোস উপস্থিত থাকে। ল্যাক্টোসের অনুপস্থিতিতে ই-কোলাই কখনোই এই এনজাইমগুলো তৈরি করেনা। প্রশ্ন হলো, কিভাবে ল্যাক্টোস উপস্থিতি বেটা-গ্যালাকটোসাইডেস ও গ্যালাকটোসাইড-পারমিয়েসে উৎপাদন নিয়ন্ত্রণ করে? ১৯৫০ সালের শুরুতে দুই ফ্রেঞ্চ বায়োলজিস্ট জ্যাক ম্যানো (Jacques Monod) ও ফ্রাঁসোয়া জ্যাকোব (François Jacob) ই-কোলাইয়ের ল্যাক্টোস মেটাবলিজমের ওপর গবেষণা শুরু করেন। তারা ই-কোলাই ব্যাকটেরিয়ার একাধিক মিউটেন্ট নিয়ে কাজ করেন এবং তিনটি সংশ্লিষ্ট জিন খুঁজে পান। বেটা-গ্যালাকটোসাইডেসের জিন LacZ, গ্যালাকটোসাইড-পারমিয়েসের জিন LacY, এবং তৃতীয় একটি জিন নাম LacI। তারা দেখতে পান LacZ বা LacY যেকোনোটির মিউটেশন ঘটলে ই-কোলাই ল্যাক্টোস হজম করতে পারেনা। কিন্তু, LacI জিনের মিউটেশনে হয় ঠিক উল্টোটি, অর্থাৎ, ল্যাক্টোস থাকুক বা না থাকুক ই-কোলাই অনবরত বেটা-গ্যালাকটোসাইডেস ও গ্যালাকটোসাইড-পারমিয়েস এনজাইমগুলো তৈরি করতে থাকে। এই ফলাফল থেকে তারা এই সিদ্ধান্তে পৌঁছেন যে, স্বাভাবিক LacI জিনের উৎপাদিত প্রোটিন কোনোভাবে LacZ ও LacY জিন দুটিকে দমিয়ে রাখে, ফলে বেটা-গ্যালাকটোসাইডেস ও গ্যালাকটোসাইড-পারমিয়েস তৈরি হতে পারে না। ল্যাক্টোস কোনোভাবে এই LacI জিনকে প্রভাবিত করে এবং দমিয়ে রাখে, ফলে LacZ ও LacY জিন দুটি ট্রান্সক্রিপ্ট হতে আর কোনো বাধা থাকে না। জ্যাকোব ও ম্যানো এই LacI জিনকে নাম দেন রিপ্রেসর (Repressor)। সাধারণ অর্থে, রিপ্রেসর হলো সেই সব জিন যারা অন্য জিনের ট্রান্সক্রিপশনে বাধা দেয়।
কিন্তু, কিভাবে ল্যাক্টোস এই LacI জিনকে প্রভাবিত করে? এই পাজেল সমাধানে সর্বপ্রথম ধারণা দেন একজন পারমাণবিক পদার্থবিদ নাম, লিও জেলার্ড (Leo Szilard)। জেলার্ড ধরণা দেন যে, ল্যাক্টোসের সাথে জোড়া লাগার কারণে LacI জিনঘটিত প্রোটিনের আকার পরিবর্তিত হয়। ফলে এই পরিবর্তিত প্রোটিন কোনো ভাবেই LacZ ও LacY জিনের রেগুলেটরি-প্রোটিন হিসেবে কাজ করতে পারে না। পরবর্তীতে ম্যানো, জ্যাকোব, এবং তাদের এক মার্কিন সহকর্মী আর্থার পারডি (Aurther Pardee) মিলে PaJaMo নামের এক ঐতিহাসিক এক্সপেরিমেন্ট চালান। এই এক্সপেরিমেন্ট জেলার্ডের ধারণাটির সত্যতা প্রমাণ করে এবং ই-কোলাইয়ের ল্যাক্টোস মেটাবলিজম সম্পর্কিত জিন নিয়ন্ত্রণ প্রক্রিয়াটি পরিষ্কারভাবে বুঝতে সাহায্য করে।
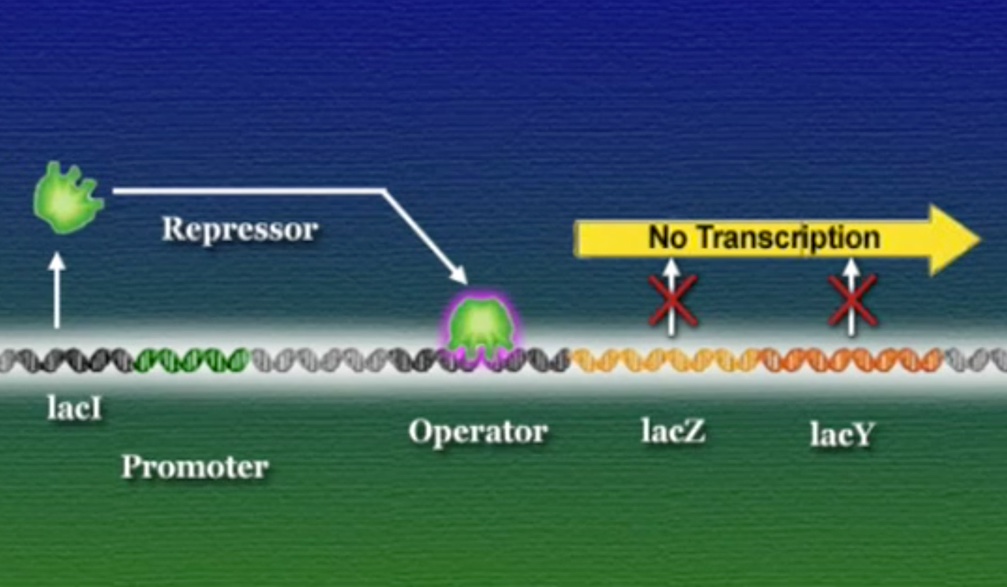
ই-কোলাই ব্যাকটেরিয়ার DNA-তে LacZ এবং LacY জিন দুটি পাশাপাশি অবস্হান করে, তারা একই রেগুলেটরি-রিজিয়ন এবং প্রমোটর ভাগাভাগি করে। ম্যানো ও জ্যাকোব এই রেগুলেটরি-রিজিয়নের নাম দেন অপারেটর (Operator)। এই অপারেটরের অধীনস্ত প্রমোটর এবং জিনগুচ্ছকে একসাথে বলে অপেরন (Operon)। অন্যদিকে, LacI একটি দূরবর্তী জিন, সুবিধার জন্য ছবিতে কাছাকাছি দেখানো হয়েছে।
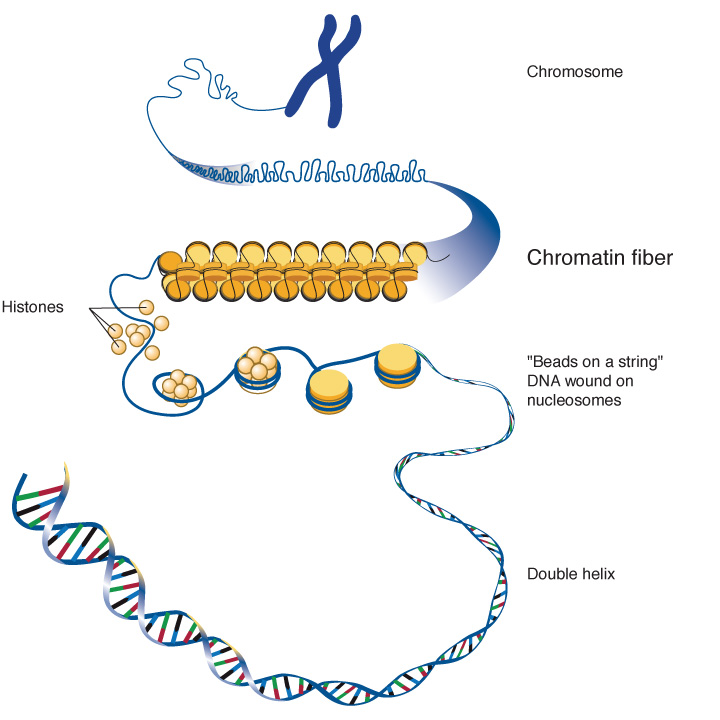
প্রোক্যারিয়টিক কোষে মাত্র একটি বৃত্তাকার ক্রোমোসোম থাকে। অন্যদিকে, ইউক্যারিয়টিক কোষে বহু সংখ্যক ক্রোমোসোমেরা আলাদা আলদাভাবে অবস্থান করে এবং এই ক্রোমোসোমগুলো মূলত প্রোটিন ও DNA দিয়ে তৈরি একটা কাঠামো। ইউক্যারিয়টিক কোষে ক্রোমোসোমের এই কাঠামোকে বলে ক্রোমাটিন (Chromatin)। ক্রোমাটিনে DNA-কে তুলোনা করা চলে ছোট ছোট রীলে পেঁচানো সুতো মতো। ক্রোমাটিনের রীলগুলো হিস্টোন প্রোটিন দিয়ে তৈরি, আর এই হিস্টোনে পেঁচিয়ে থাকে DNA। ক্রোমাটিনের দৈর্ঘ্য ও কাঠামোর জন্য ইউক্যারিয়টিক কোষে জিন নিয়ন্ত্রণ পদ্ধতিও ভিন্ন।
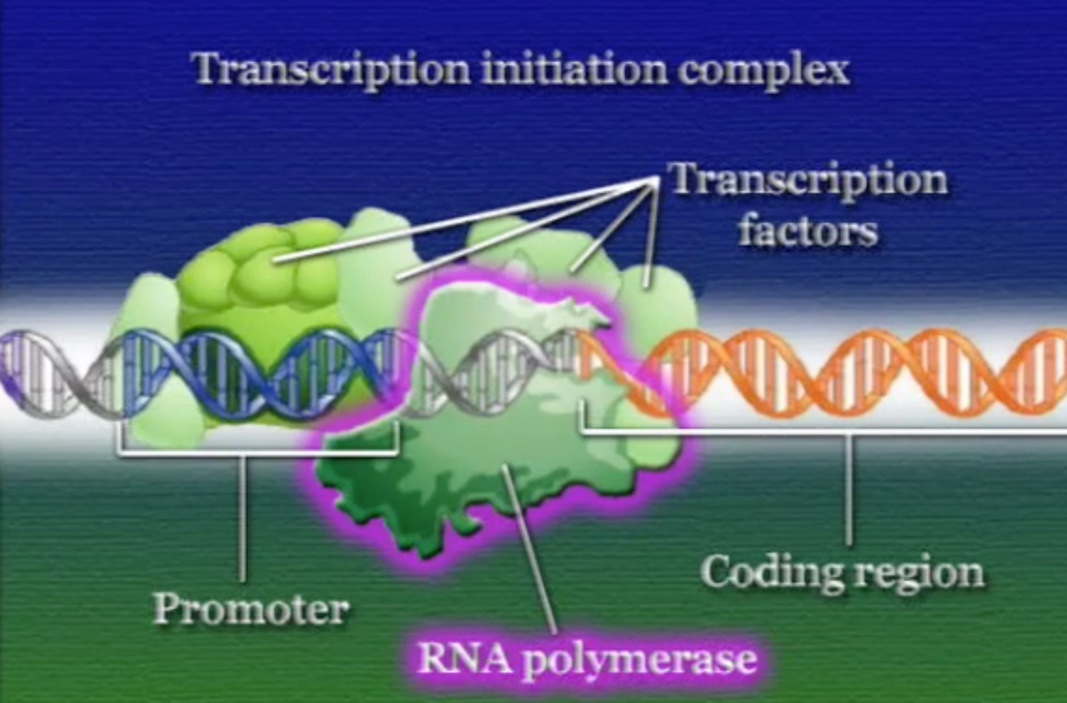
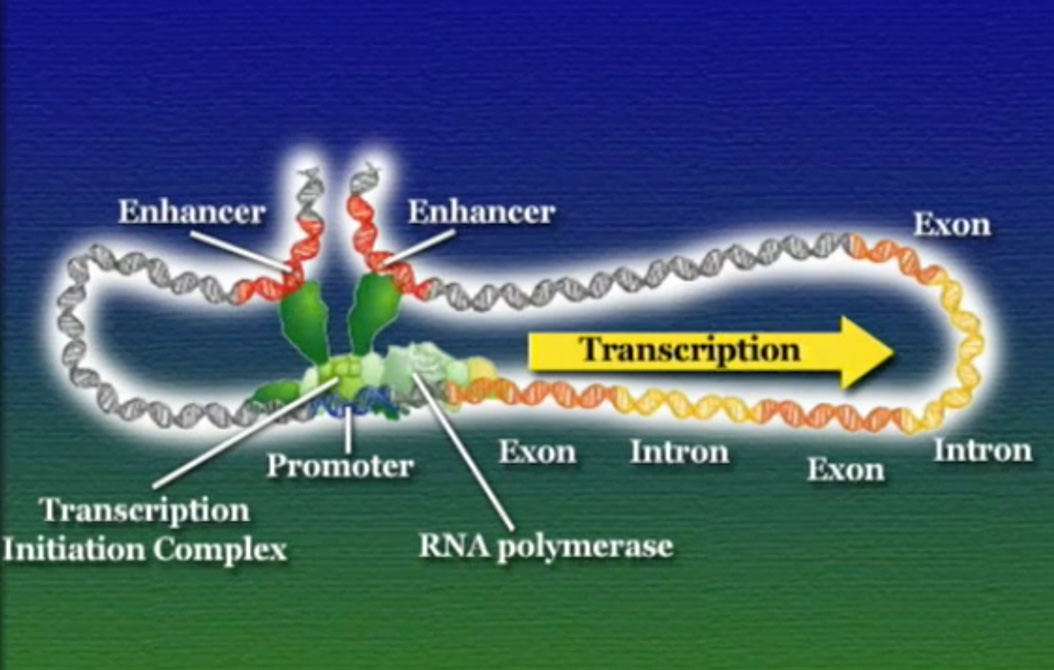
ইউক্যারিয়টিক কোষে প্রতিটি জিনের নিজস্ব অপেরন থাকে, অর্থাৎ, ইউক্যারিয়টিক কোষে জিনেরা কখনো গুচ্ছ অবস্হায় থাকে না। জিনের প্রমোটর অংশে কিছু সাধারণ (Generic) প্রোটিন আটকে থাকে যারা RNA-পলিমারেসকে ট্রান্সক্রিপ্টশনে সাহায্য করে কিন্তু জিন নিয়ন্ত্রণ করে না। এই সাহায্যকারী প্রোটিনদের বলে ব্যাসেল-ট্রান্সক্রিপশন-ফেক্টর (Basal transcription factor) এবং এরা যেকোনো জিনের প্রমোটরে সাথে যুক্ত হতে পারে। অন্যদিকে, রেগুলেটরি-রিজিয়ন একই প্রমোটরে আরো কিছু প্রোটিন যোগ করে, এই রেগুলেটরি-প্রোটিনগুলোকে বলে রেগুলেটরি-ট্রান্সক্রিপশন-ফেক্টর, এরা জিন নিয়ন্ত্রণের জন্য দায়ী, এবং এরা শুধুমাত্র একটি নির্দিষ্ট জিনের প্রমোটরে সাথে যুক্ত হতে পারে। রেগুলেটরি এবং ব্যাসেল-ট্রান্সক্রিপশন-ফেক্টরের এই বিশাল প্রোটিন বাহিনীকে একসাথে বলে ট্রান্সক্রিপশন-ইনিশিয়েশন-কমপ্লেক্স (Transcription initiation complex)। এই ট্রান্সক্রিপশন-ইনিশিয়েশন-কমপ্লেক্স কাঠামোর কারণেই RNA-পলিমারেস জিনের প্রমোটর অঞ্চলটি খুঁজে পায়। ট্রান্সক্রিপশনের সময় RNA-পলিমারেস পুরো ট্রান্সক্রিপশন-ইনিশিয়েশন-কমপ্লেক্সকে নিয়ে সামনে এগুতে থাকে। অনেকটা রথযাত্রার মতো।
প্রোক্যারিয়টিক জিনের সাথে ইউক্যারিয়টিক জিনের একটা বড় পার্থক্য হলো, ইউক্যারিয়টিক জিনে একাধিক রেগুলেটরি-রিজিয়ন থাকে এবং এরা জিন থেকে বহু দূরে ছড়িয়ে ছিটিয়ে থাকতে পারে। এই দূরবর্তী রেগুলেটরি-রিজিয়নগুলোকে বলে এনহেন্সার (Enhencer)। প্রশ্ন হলো, এই এনহেন্সারগুলো কিভাবে তাদের দূরবর্তী জিনকে নিয়ন্ত্রণ করে? ট্রান্সক্রিপশন শুরুর আগে DNA এমনভাবে বেঁকে যায় যে, একাধিক এনহেন্সার তাদের সংশ্লিষ্ট প্রোমোটরের কাছাকাছি চলে আসে এবং প্রোমোটরে রেগুলেটরি-প্রোটিন আটকাতে সাহায্য করে।
ক্রোমাটিনের হিস্টোন প্রোটিন নিজেও জিন নিয়ত্রণে অবদান রাখে। যেমন, ট্রান্সক্রিপশন শুরুর আগে হিস্টোন অনেকটা রীল থেকে সুতোর পাক খোলার মতো জিনকে আলগা করে দেয়, ফলে RNA-পলিমারেস তার মহাযজ্ঞ শুরু করতে পারে। আর যে জিনগুলো হিস্টোনের প্যাকেজিং ভেতর চাপা থাকে RNA-পলিমারেস তাদের খুঁজেই পায়না।
ইউক্যারিয়টিক জিন নিয়ত্রণ একটা বিশাল বিষয়। এখানে শুধু প্রাথমিক ধারণা দেয়া হয়েছে মাত্র।
অবলম্বনে: “The Great Courses” থেকে প্রকাশিত “Biology: The Science of Life” by “Stephen Nowicki”