একটি গ্লুকোস অণু থেকে শক্তি পাওয়াকে শুধু এক লাইনের একটি সমীকরণের মাধ্যমে লিখা যায়।

সমীকরণের বাঁয়ের গ্লুকোস হলো কার্বনের রিডিউসড (Reduced) অবস্থা, কারণ, গ্লুকোসে কার্বন তার প্রতিটি ইলেক্ট্রন নিজের কাছে রেখে দেয়। আর, সমীকরণের ডানের কার্বন-ডাই-অক্সাইড কার্বনের সবচেয়ে অক্সিডাইজড (Oxidized) অবস্থা, কারণ কার্বন-ডাই-অক্সাইডে কার্বনের শেষ কক্ষপথের ইলেট্রনগুলো মোটামুটি অক্সিজেনের দখলে চলে যায়। সমীকরণটি একটি রিডক্স বিক্রিয়া, যেখানে কার্বনের সম্পূর্ণ অক্সিডেশন ঘটছে। সমস্যা হলো, এই রিডাক্স বিক্রিয়াটি যদি সমীকরণটির মতো এক ধাপে ঘটে তবে, হটাৎ করেই যে শক্তি উৎপন্ন হবে সেটা কোষের জন্য অত্যন্ত বিপদজনক। তাই, কোষ সমীকরণটি তিনটি ক্যাটাবোলিক পাথওয়ের মাধ্যমে সমাধান করে। এই পাথওয়েগুলোর নাম হলো, গ্লাইকোলিসিস (Glycolysis) বা গ্লাইকোল্যাটিক-পাথওয়ে, ক্রেব-সাইকেল (Krebs cycle), এবং ETC বা ইলেক্ট্রন-ট্রান্সপোর্ট-চেইন (Electron transport chain)। এদের এক সাথে বলে, সেলুলার-রেস্পিরেশন (Cellular respiration)।

প্রথমেই তিনটির মধ্যে কিছু পার্থক্য দেয়া যাক। গ্লাইকোলিসিস ঘটে কোষের সাইটোপ্লাসমে (Cytoplasm), অন্যদিকে ক্রেব-সাইকেল এবং ETC ঘটে মাইটোক্রোন্ডিয়ায় (mitochondria)। গ্লাইকোলিসিস ঘটে অক্সিজেনের অনুপস্থিতিতে তাই একে বলে এনারোবিক-রেস্পিরেশন (Anaerobic respiration)। ওদিকে, ক্রেব-সাইকেল এবং ETC-এর জন্য অক্সিজেন অপরিহার্য, তাই এদের বলে এরোবিক-রেস্পিরেশন (Aerobic respiration)। গ্লাইকোলিসিস এবং ক্রেব-সাইকেল দুটোই হচ্ছে সাবস্ট্রেট-লেভেল-ফসফোরেলেশন (Substrate level phophorylation) ধরণের। সাবস্ট্রেট শব্দটা এনজাইমের সাথে সম্পর্কযুক্ত। তাই, সাবস্ট্রেট-লেভেল-ফসফোরেলেশন কথাটার মানে হচ্ছে, পাথওয়ের প্রতিটি রিডাক্স বিক্রিয়াতেই কাপলিং-এনজাইম ব্যবহার করা হয়। আর, ETC হলো অক্সিডেটিভ-ফসফোরেলেশন (Oxidative-phophorylation)। ETC-তে কোনো কাপলিং-এনজাইম ব্যবহার করা হয় না। ETC-এর রিডাক্স বিক্রিয়া কাপলিং-এনজাইম ছাড়াই পরোক্ষভাবে ATP রিচার্জ করে।

গ্লাইকোলিসিসের রয়েছে মোট ১০টি রিডাক্স বিক্রিয়া। এই পাথওয়ের শুরু হয় ছয়-কার্বনযুক্ত একটি গ্লুকোস থেকে আর শেষ হয় তিন-কার্বনযুক্ত দুটি পাইরুভেট (Pyruvate) অণুতে। গ্লাইকোলিসিসের প্রথম ৫টি বিক্রিয়াকে বলে এনাৰ্জি-ইনভেস্টমেন্ট-ফেজ (Energy investment phase)। এই ধাপে গ্লুকোস রূপান্তরিত হয় গ্রিস্যারেলডিহাইড-৩-ফসফেট বা জি৩পি (Glyceraldehyde-3-phosphate) -তে। ধাপটি এন্ডোর্গনিক বা শক্তি-শোষক, এতে দুটি ATP খরচ হয়। ৬নম্বর থেকে শেষপর্যন্ত বিক্রিয়াগুলোকে বলে এনাৰ্জি-হার্ভেস্টিং-ফেজ (Energy harvesting phase)। ধাপটি এক্সার্গনিক বা শক্তি-দায়ক। এই ধাপে, দুটি NAD+ কে রিডাকশন বিক্রিয়ার মাধ্যমে NADH-এ পরিণত করা হয় এবং সেই সাথে চারটি ATP-কে রিচার্জ করে হয়। গ্লাইকোল্যাটিক-পাথওয়ের নেট অর্জন হলো দুটি ATP এবং দুটি NADH। গ্লাইকোলিসিসের সীমাবদ্ধতা হলো, কোষে NAD+ এর পর্যাপ্ততা। কোনো কারণে যদি কোষে পর্যাপ্ত NAD+ না থাকে তবে গ্লাইকোলিসিস ৬নম্বর বিক্রিয়ায় এসে থেমে যাবে। ফলে, পুরো পাথওয়ে বন্ধ হয়ে যাবার সম্ভাবনা তৈরি হয়। এই অবস্থায় কোষ ফার্মেন্টেশন (Fermentation) প্রক্রিয়ায় NADH-এর অক্সিডেশন ঘটিয়ে NAD+ পরিমাণ বৃদ্ধি করে। কোষ ভেদে এই ফার্মেন্টেশন দুই রকমের, এলকোহোল-ফার্মেন্টেশন (Alcohol fermentation) এবং ল্যাকটিক-অ্যাসিড-ফার্মেন্টেশন (Lactic acid fermentation)।

ঈস্ট (Yeast) এলকোহোল-ফার্মেন্টেশন ঘটায়। এরা প্রথমে পাইরুভেটকে থেকে অ্যাসিটালডিহাইড (Accetaldehyde), এরপর অ্যাসিটালডিহাইডকে রিডিউসড করে ইথানল তৈরি করে। অ্যাসিটালডিহাইডকে রিডিউসড করার সময় NADH অক্সিডাইজড হয়ে NAD+ এ পরিণত হয়।

ল্যাকটিক-অ্যাসিড-ফার্মেন্টেশন আমাদের কাছে অত্যন্ত বসুপরিচিত। কিছু কিছু ব্যাকটেরিয়া এই ফার্মেন্টেশনের মাধ্যমে দুধ থেকে পনির এবং দই তৈরি করে। একটানা পরিশ্রমে আমাদের মাংসপেশীতে অক্সিজেনের অভাব ঘটে। তখন আমাদের পেশীকোষ পাইরুভেটকে সরাসরি রিডিউসড করে ল্যাকটিক-অ্যাসিডে পরিণত করে। মাংসপেশীতে এই ল্যাকটিক-অ্যাসিডের কারণে আমরা পেশীতে ব্যাথা অনুভব করি। এই ল্যাকটিক-অ্যাসিড পরবর্তীতে যকৃতে আবার পাইরুভেটে রূপান্তরিত হয়। ল্যাকটিক-অ্যাসিড-ফার্মেন্টেশনে পাইরুভেট রিডিউসড হবার সাথে সাথে NADH অক্সিডাইজড হয়ে NAD+ এ পরিণত হয়।
গ্লাইকোলিসিস পাথওয়ে গ্লুকোসের মাত্র ৩% শক্তি বের করে আনে। যদিও পাইরুভেট গ্লুকোস থেকে অপেক্ষাকৃত কম রিডিউসড, এই পাইরুভেটে প্রচুর শক্তি রয়ে যায়। পাইরুভেটকে পরবর্তী প্রক্রিয়াজাতকরণের জন্য পাঠিয়ে দেয়া হয় মাইটোক্রোন্ডিয়ার ম্যাট্রিক্সে, যেখানে ক্রেব-সাইকেল পাইরুভেটকে ভেঙ্গে কার্বনের সবচেয়ে অক্সিডাইজড অবস্থা কার্বন-ডাই-অক্সাইডে পরিণত করে। জার্মানির স্যার হ্যান্স ক্রেব (Sir Hans Krebs) ১৯৩০ সালের দিকে ক্রেব-সাইকেল পাথওয়ে প্রস্তাব করেন। ১৯৫৩ সালে এইজন্য উনি নোবেল পদক পান। ক্রেব-সাইকেলে রয়েছে ৮টি রিডাক্স বিক্রিয়া। ৮নম্বর বিক্রিয়ার উৎপাদিত বস্তু আবার ১নম্বর বিক্রিয়ায় ব্যবহার করা হয়, তাই এই ক্রেব-সাইকেল চক্রাকারে ৯টি ধাপে সম্পন্ন হয়। ক্রেব-সাইকেলের প্রস্তুতি শুরু হয় সাইটোপ্লাসম থেকে।

ক্রেব-সাইকেলে ঢোকার আগে পাইরুভেটকে অক্সিডাইজড করে প্রায় ৬০টি প্রোটিনের বিশাল একটা এনজাইম-যন্ত্র, নাম পাইরুভেট-ডিহাইড্রোজেনাস-কমপ্লেক্স (Pyruvate dehydrogenase complex)। এটি একটি শক্তিশালী এক্সার্গনিক রিডাক্স বিক্রিয়া। এতে অক্সিডাইজ হয়ে পাইরুভেটের তিন কার্বনের একটি কার্বন-ডাই-অক্সাইড আকারে বেরিয়ে যায়, বাকি দুটি কার্বন মিলে তৈরি করে অ্যাসিটালডিহাইড। বিক্রিয়া থেকে নির্গত শক্তি থেকে এই অ্যাসিটালডিহাইড যুক্ত হয় কোএনজাইম-এ (Coenzyme A) নামক এক প্রোটিনের সাথে, ফলে তৈরি হয় অ্যাসিটাইল-কোএ (Acetyl CoA)। ফসফোয়নহাইড্রাইডের মতোই অ্যাসিটালডিহাইড এবং কোএনজাইম-এ দুটির বন্ধন অত্যন্ত শক্তিশালী। এরপর এই অ্যাসিটাইল-কোএ কে নিয়ে যাওয়া হয় মাইটোক্রোন্ডিয়ার ম্যাট্রিক্সে।
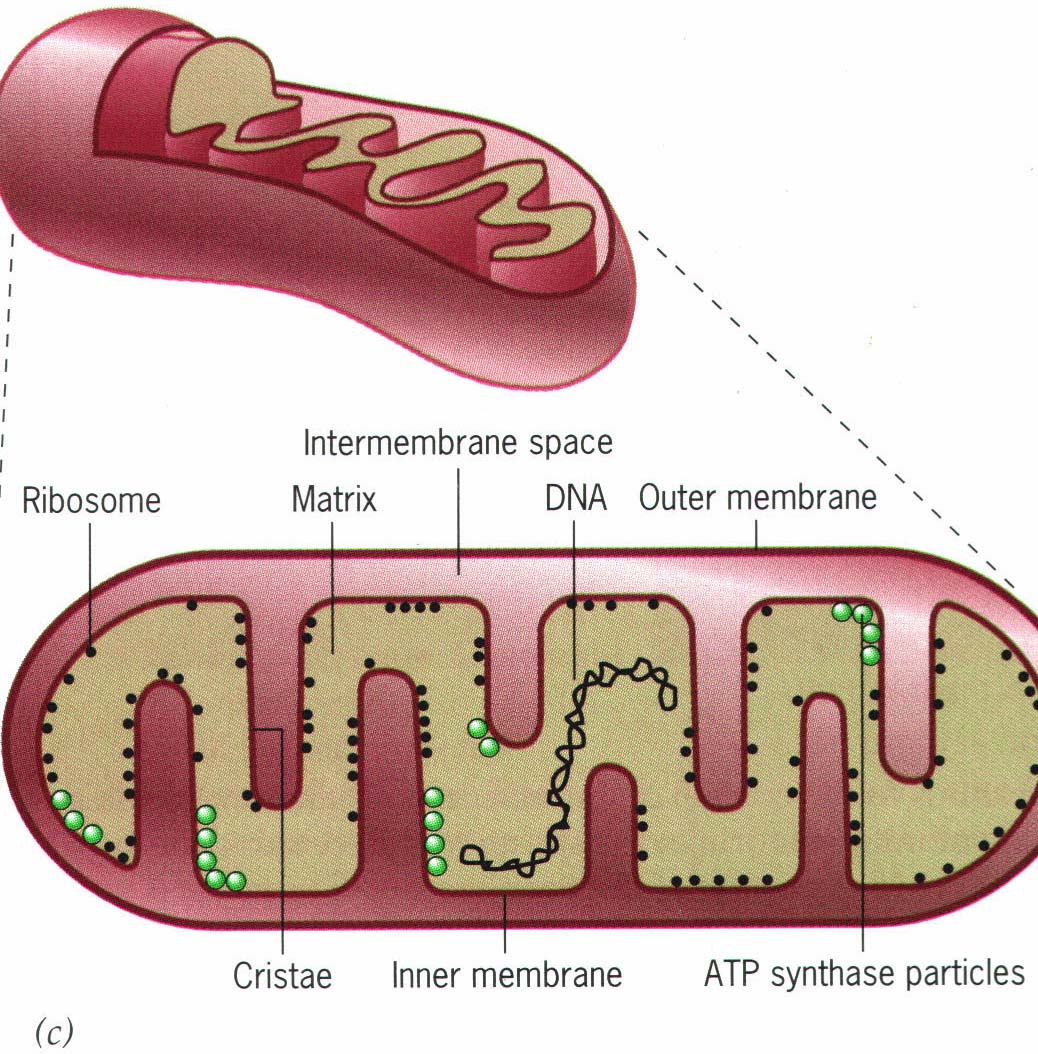
প্রাণী কিংবা উদ্ভিদ প্রতিটি ইউক্যারিয়টিক কোষেই মাইটোক্রোন্ডিয়া পাওয়া যায়। এই মাইটোক্রোন্ডিয়ার দুটি মেমব্রেন। মেমব্রেন দুটির মাঝের ফাঁকা অংশকে বলে ইন্টারমেমব্রেন-স্পেস (Intermembrane space)। ভেতরের মেমব্রেনটি আঁকাবাঁকা হয়ে হাতের আঙ্গুলের মতো ভাঁজ হয় থাকে। এই ভাঁজগুলোকে বলে ক্রিস্টি (Cristae)। আর এই ক্রিস্টির ভেতরের অংশটিকে বলে ম্যাট্রিক্স (Matrix)। ম্যাট্রিক্স মাইটোক্রোন্ডিয়ায় সবচেয়ে ভেতরের অংশ।
ক্রেব-সাইকেলের অ্যাসিটাইল-কোএ প্রথম ধাপে অক্সালোঅ্যাসিটেটের (Oxaloacetate) সাথে যুক্ত হয়ে সাইট্রেট (Citrate) তৈরি করে। এই প্রক্রিয়ায় অ্যাসিটাইল-কোএ তার কোএনজাইম-এ অংশটি হারায়। এই কোএনজাইম-এ টি ক্রেব-সাইকেলের ৫ এবং ৬নম্বর ধাপে পুনঃব্যবহার হয়। ৪ এবং ৫নম্বর ধাপে গ্লুকোস তার বাকি সমস্ত কার্বনকে দুটি কার্বন-ডাই-অক্সাইড আকারে হারিয়ে ফেলে। ৫নম্বর ধাপে ATP রিচার্জ করা হয়, তবে সরাসরি নয়। প্রথমে GDP বা গুয়ানোসিন-ডাইফসফেটকে (Guanosine diphosphate) ফসফোরেলেশনের মাধ্যমে GTP-তে রূপান্তর করা হয়। GTP আসলে তিনটি ফসফেট-গ্রুপযুক্ত গুয়ানিন নিউক্লিইক-অ্যাসিড। এই GTP পরবর্তীতে ADP কে ফসফোরেলেশনের মাধ্যমে ATP-তে রূপান্তর করে। ৪, ৫, ৭, এবং ৯নম্বর ধাপে NAD+ এবং FAD+ দের রিডিউসড করে যথাক্রমে NADH এবং FADH2-তে পরিণত করা হয়। ক্রেব-সাইকেলের ৮নম্বর এবং শেষ ধাপের চূড়ান্ত বস্তু অক্সালোঅ্যাসিটেট, যেটা পুনরায় ১নম্বর ধাপে ফিরে যায়। আর এভাবেই চক্রাকারে চলতে থাকে ক্রেব-সাইকেল।

গ্লাইকোলিসিস এবং ক্রেব-সাইকেল শেষে যদি হিসেব করা হয় তবে আমরা পাবো নেট ৪টি ATP। দুটি গ্লাইকোলিসিস থেকে, বাকি দুটি ক্রেব-সাইকেল থেকে। অথচ, এটা একটি গ্লুকোস অণু মাত্র ৬% শক্তি। এবং, ক্রেব-সাইকেল শেষ গ্লুকোস পুরোপুরি অক্সিডাইজ হয়ে কার্বন-ডাই-অক্সাইডে পরিণত হয়। তাহলে, বাকি শক্তি গেলো কোথায়? আমরা এই পাথওয়ে দুটি থেকে কিছু NADH এবং FADH2 পেয়েছিলাম, এবার তাদের হিসাব করা যাক। ক্রেব-সাইকেল শেষে আমরা এক অণু গ্লুকোস থেকে মোট ১২টি NADH এবং FADH2 পাই। গ্লাইকোলিসিসে ২টি NADH, ক্রেব-সাইকেল প্রস্তুতিতে ২টি NADH, এবং ক্রেব-সাইকেল থেকে ৬টি NADH ও ২টি FADH2। কোষ এই NADH ও FADH2-গুলোকে সরাসরি ATP-এর মতো শক্তির উৎস হিসেবে ব্যবহার করতে পারেনা। কোষ এদের ব্যবহার করে পরবর্তী পাথওয়ে ETC তে।

ETC পাথওয়ে কিভাবে ATP রিচার্জ করে সেই সম্পর্কে সর্বপ্রথম ধারণা দেন ইংল্যান্ডের পিটার মিশেল (Peter Mitchell ) ১৯৬১ সালে। তার এই ধারণাকে বলে কেমিওসমোসিস তত্ত্ব (Chemiosmosis theory)। পরবর্তীতে পরীক্ষণের মাধ্যমে ETC পাথওয়ের কেমিওসমোসিস তত্ত্বটি বহুলাংশে প্রতিষ্ঠিত হয়। এই তত্ত্বের জন্য মিশেল ১৯৭৮ সালে নোবেল পদক পান। মাইটোক্রোন্ডিয়ার ভেতরের মেমব্রেনে বিশেষ ধরণের প্রচুর আয়ন-চ্যানেল রয়েছে। এই আয়ন-চ্যানেলগুলো গুচ্ছ বা কমপ্লেক্স (Complex) আকারে কাজ করে। প্রতিটি কমপ্লেক্সের ভিন্ন ভিন্ন নাম আছে। এই আয়ন-চ্যানেলগুলো মূলত এক্টিভ-আয়ন-চ্যানেল। সক্রিয় অবস্থায় চ্যানেলগুলো হাইড্রজেন-আয়ন বা প্রোটনকে (H+) ম্যাট্রিক্স থেকে ইন্টারমেমব্রেন-স্পেসে বের করে দেয়। তাই, এই আয়ন-চ্যানেলগুলোকে বলা হয় প্রোটন-পাম্প।

গ্লাইকোলিসিস এবং ক্রেব-সাইকেলে পাওয়া NADH এবং FADH2-দের বহন করা ইলেক্ট্রনগুলো এই প্রোটন-পাম্পের কমপ্লেক্সগুলোতে রিডাক্স বিক্রিয়া চালাতে সহায়তা করে। আর, এই রিডক্স সক্রিয় করে তোলে প্রোটন-পাম্পগুলোকে। কমপ্লেক্সগুলোতে ইলেক্ট্রনগুলো ধাপে ধাপে শক্তি হারাতে থাকে। এইজন্যই ETC-কে বলে অক্সিডেশন-ফসফোরেলেশন। এই রিডাক্স বিক্রিয়ায় কোনো এনজাইম দরকার হয় না। প্রোটনকে ম্যাট্রিক্স থেকে বের করে দেবার কারণে, ম্যাট্রিক্স এবং ইন্টারমেমব্রেন-স্পেসের মধ্যে প্রোটনের ঘনত্ব ভিন্ন হয়। ম্যাট্রিক্সে ঘনত্ব কম, ইন্টারমেমব্রেন-স্পেসে বেশি। প্রোটন ঘনত্বের এই তারতম্যকে বলে প্রোটন-গ্র্যাডিয়ান্ট (Proton gradient)। এই রিডাক্স বিক্রিয়ার চুড়ান্ত বস্তু হলো অক্সিজেন অণু, এই অক্সিজেন পরবর্তীতে প্রোটন বা হাইড্রোজেন-আয়নের সাথে যুক্ত হয়ে পানি তৈরি করে।

প্রোটন-গ্র্যাডিয়ান্ট ব্যবহার করে ATP রিচার্জ করার জন্য দায়ী আরেক ধরণের মেমব্রেন-বাউন্ড প্রোটিন, নাম ATP-সিন্থেস (ATP synthase)। ATP-সিন্থেসকে তুলোনা করা চলে বিদ্যুৎ উৎপাদন কেন্দ্রের টার্বাইনের সাথে। এর তিনটি অংশ আছে। রোটর (Roter), রড (Rod), এবং নব (Knob)। উচ্চ ঘনত্বের ইন্টারমেমব্রেন-স্পেস থেকে প্রোটন স্রোতের মতো ATP-সিন্থেসের মধ্যেদিয়ে ম্যাট্রিক্সে ঢুকতে থাকে, ফলে প্রোটিনের রোটর ও রড অংশদুটো টার্বাইনের মতো ঘুরতে শুরু করে। এক পরীক্ষণ অনুসারে এই ঘূর্ণনগতি প্রতি মিনিটে প্রায় ৫০ বার বা ৫০RPM (Revolutions per minute)। আর এই যান্ত্রিক শক্তি ব্যবহার করেই ATP-সিন্থেস তার নব অংশে ADP ফসফোরেলেশনের মাধ্যমে ATP রিচার্জ করে।

আমরা জীবনধারণের জন্য সরাসরি গ্লুকোস খাই না। বরং, আমাদের খাবার মূলত আমিষ (Protein), শর্করা (Carbohydrate), চর্বি ( Fat)। সেলুলার-রেস্পিরেশন হলো শরীরের মেটাবোলিক পাথওয়ের একেবারেই কেন্দ্রীয় অংশ। শরীরের অন্যান্য কিছু মেটাবোলিক পাথওয়ে খাবারের বিভিন্ন উপাদানগুলোকে ভেঙ্গে গ্লাইকোলিসিস এবং ক্রেব-সাইকেলের উপযোগী করে তোলে। আর, এই ভেঙ্গে ফেলা অপেক্ষাকৃত সরল উপাদানগুলো পরবর্তীতে সেলুলার-রেস্পিরেশন পাথওয়েগুলোতে পাঠিয়ে দেয়া হয়।
এবার আসা যাক চূড়ান্ত হিসেবে, ETC শেষে কোষ মোট ৩৮টি ATP রিচার্জ করে। ETC একাই রিচার্জ করে ৩৪টি ATP। সেলুলার-রেস্পিরেশন শেষে কোষ এক অণু গ্লুকোসের প্রায় ৬০% শক্তি কাজে লাগায়। মনে হতে পারে, সেলুলার-রেস্পিরেশনে গ্লুকোসের মোট শক্তির একটি বিশাল অংশ, প্রায় ৪০% অপচয় হয়। ভালো কথা, কিন্তু, এটাও মনে রাখা দরকার, মানুষের তৈরি সবচেয়ে কর্মদক্ষ জ্বালানি-বান্ধব ইঞ্জিন তার জৈব-জ্বালানি মাত্র ২৫% কাজে লাগায়।
অবলম্বনে: “The Great Courses” থেকে প্রকাশিত “Biology: The Science of Life” by “Stephen Nowicki”